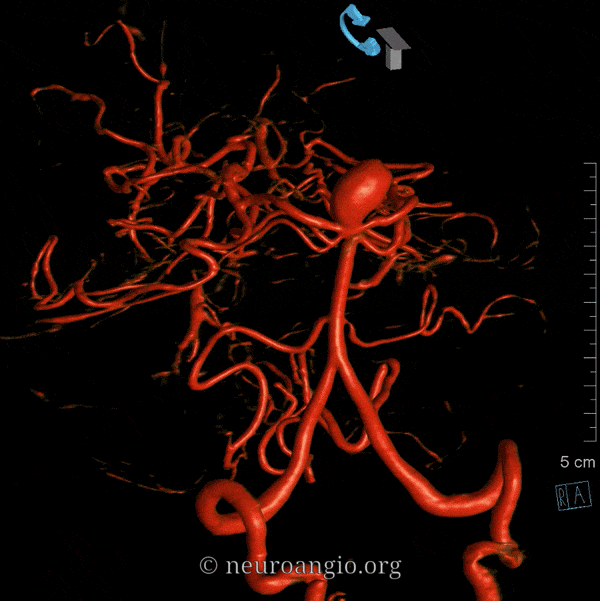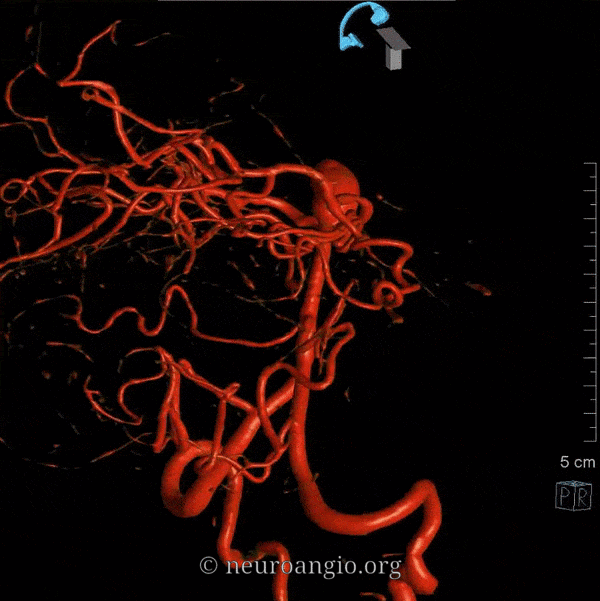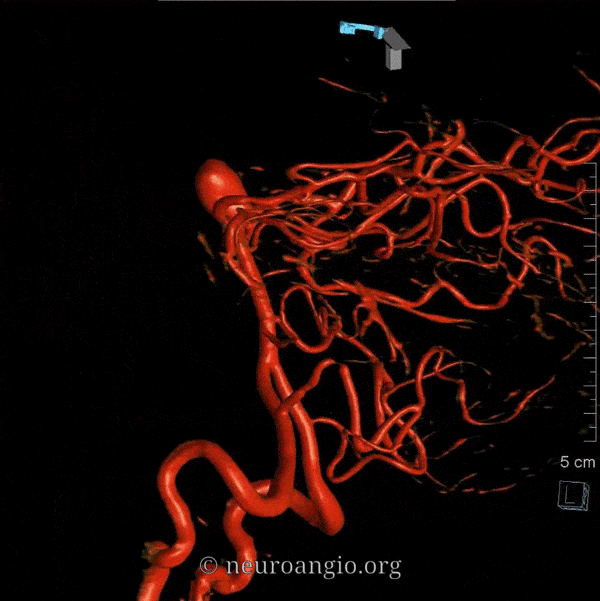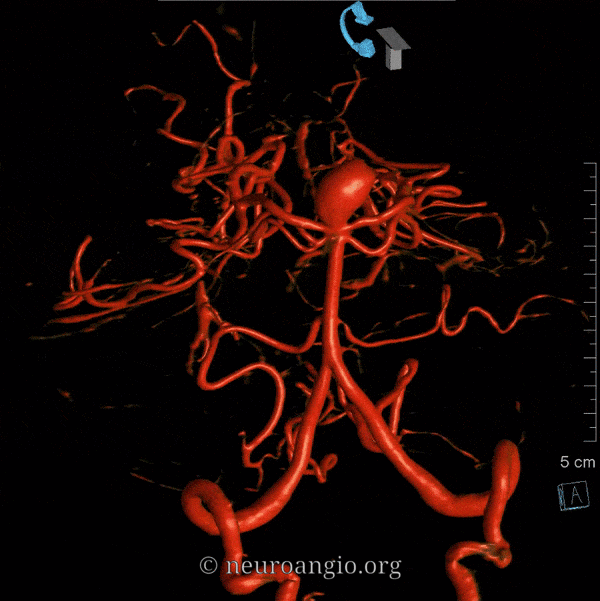
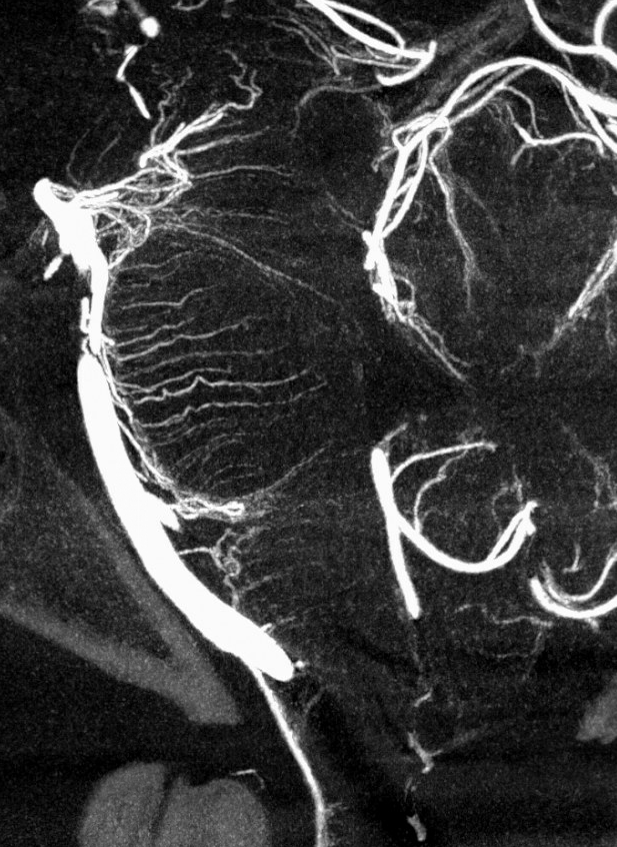
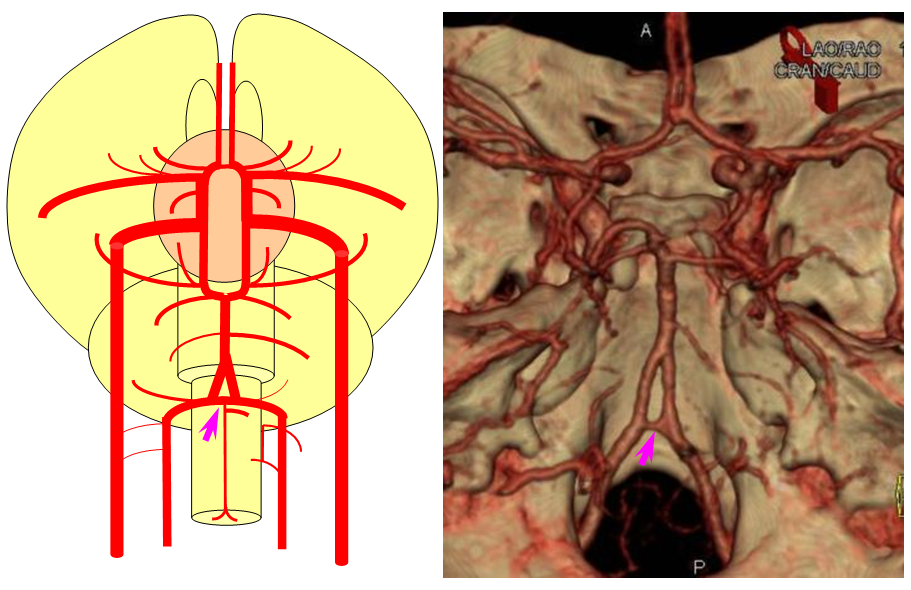
The basilar artery is formed by fusion of the longitudinal neural system, which in its most primitive form consists of loosely connected channels running along the undersurface of the brainstem. Lasjaunias and his collegues view arterial system of the brainstem and cerebellum as a natural extension of the segmental arrangement found in the spinal cord. The conceptual brilliance of this view allows one to understand all the myriad variations to which the basilar artery and its daughter vessels are subjected. In other words, if you consider the basilar artery to be a continuation of the anterior spinal artery, and its named branches and perforators as homologs of the coronary and sulco-comissural arteries (see Spinal Vascular Anatomy section), then the overall arrangement and its possible variations make perfect sense. The following diagrams serve to illustrate this concept. On a personal note, I generally find anatomical diagrams to be at least somewhat wanting; when applied to the living body, they too often suffer from both rigidity and inconsistency, and almost universally fall short of the predictive potential for which their creation was originally intended. In this case, however, I believe that the genius of Lasjaunias (and supporting giants), may prove an exception. It is not, by any stretch, The Periodic Table, but some time investment into a bit of theoretical discussion is likely to produce major dividends. Below is a diagram of cervical spinal vasculature (left), and brainstem vasculature (right), without the cerebellum.
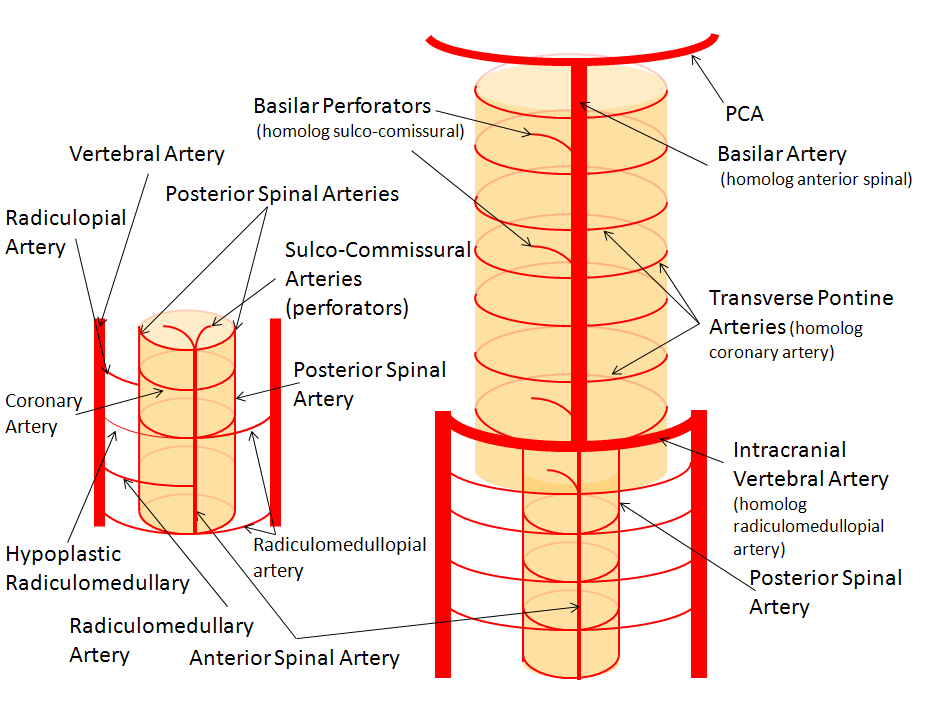
The image on the LEFT represents cervical spinal cord arterial supply, which consists of the anterior spinal artery and a paired, loose network of posterolateral vessels known as the posterior spinal arteries, and which are conceptually represented here as contiguous vessels (which is at least mostly true in the cervical spine). The anterior and posterior spinal systems are connected by anastomoses running along the circumference of the cord, although known as “coronary” arteries, are conceptually quite clear. A number of perforating arteries into the substance of the cord exist; when arising from the anterior spinal artery and penetrating through the ventral cord sulcus, they are named “sulco-comissural” arteries. The entire spinal cord system is supplied via segmental radiculomedullary arteries, which connect the vertebral artery to the anterior spinal artery. In practice, as you know, the radiculomedullary and radiculopial arteries are fewer, and may arise from longitudinal vessels other than the vert. Radiculopial arteries are those which supply the posterior spinal system. Radiculomedullopial arteries are those which happen to supply both anterior and posterior spinal systems simultaneously, sometimes via a coronary artery, and at other times via separate connections to the posterior spinal system. For a more complete discussion of spinal vasculature, see Spinal Vascular Anatomy section, particularly Spinal Arterial Anatomy.Now, lets add the brainstem to the spinal cord, and use existing arterial vascular networks to furnish its supply. Think of the brainstem as just a somewhat larger diameter biomass than the spinal cord, and things start to make sense. The unapaired basilar artery is a homolog of the equally unpaired anterior spinal artery. The intracranial vertebral arteries, although obliquely oriented, are essentially homologs of the radiculo-medullary arteries, inasmuch as they serve as trasverse connections between the extraspinal vertebral system and the anterior spinal axis. The transverse pontine arteries are homologs of the coronary arteries. The basilar perforators are homologs of the sulco-comissural arteries. The posterior spinal arteries, in the superior cervical spine, are sometimes termed “Lateral spinal arteries”. This creates much unnecessary confusion, but the posterior spinal system and lateral spinal system are one and the same longitudinal arrangement.
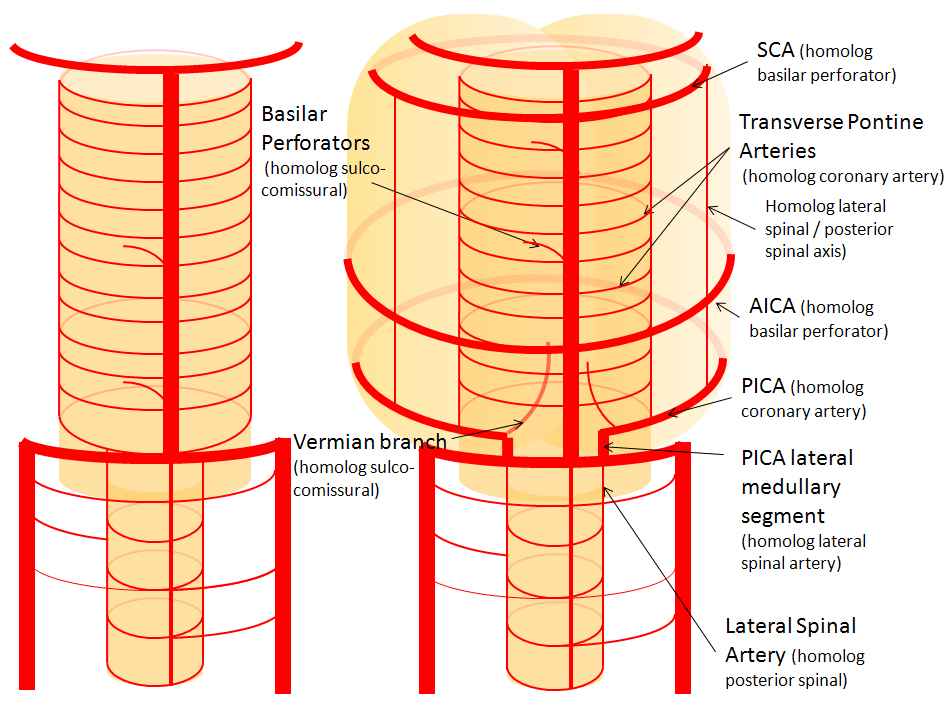
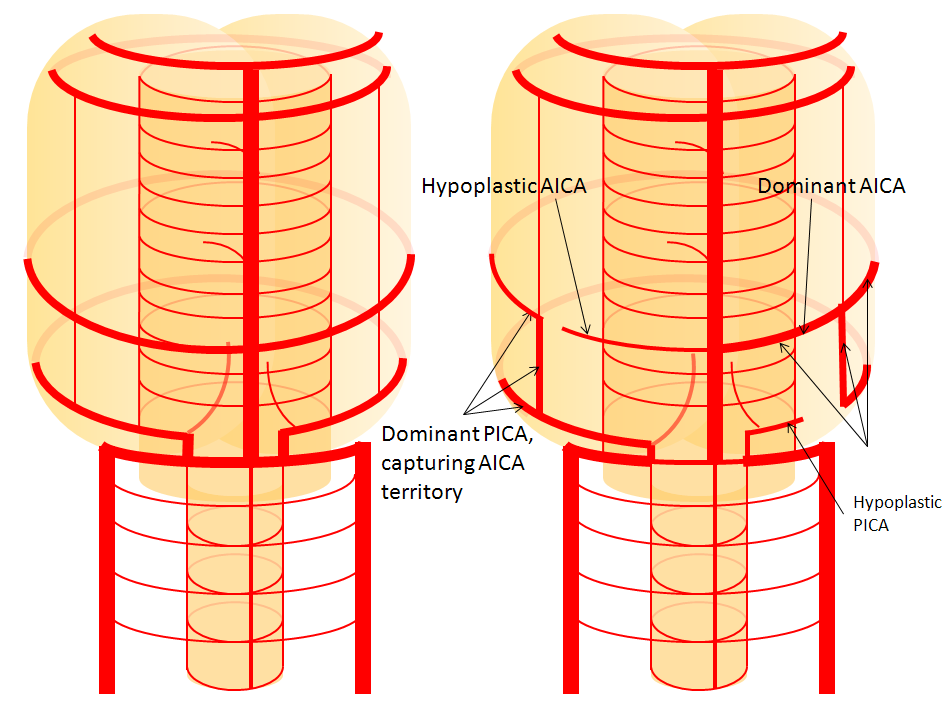
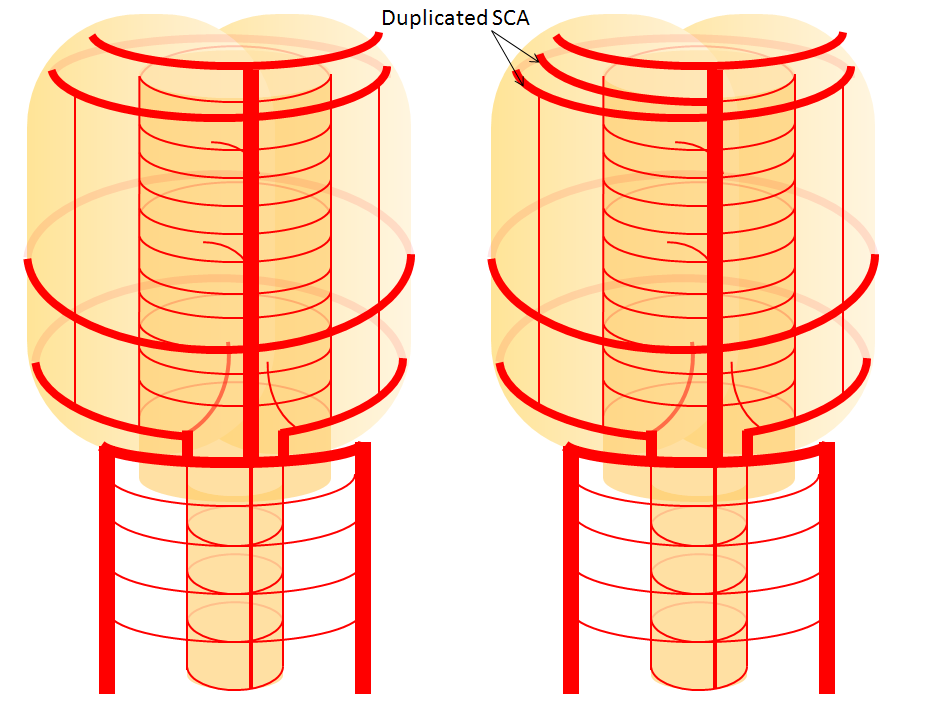
Now, add a cerebellum to the back of the brainstem, — again, simply more biomass — and use existing arteries to supply it. As the cerebellum develops, some of the transverse pontine perforators are recruited to capture the cerebellar hemispheric territory. Superiorly, this happens relatively consistently, and produces what is known as the Superior Cerebellar Artery. At the mid to lower basilar segment, a homologous enlarging channel is the AICA. At the bottom, the Posterior Inferior Cerebellar Artery (PICA) is the latest addition to cerebellar supply, Unlike AICA and SCA, it seems to arise from the lateral spinal system (yet nervertheless also a coronary artery homolog). The vermian arteries (of which only inferior is shown here) may be regarded as homologs of the sulco-comissural vessels. There are many advantages to viewing the vertebrobasilar system in this way. All kinds of variants become quite predictable. For example, duplicated and triplicated SCAs and AICAs are simply persistence of adjacent transverse pontine (or midbrain) arteries in supply of the cerebellar hemisphere. AICAs arising higher or lower along the basilar are either results of dominance of higher or lower transverse arteries, or consequent to a relatively “short” basilar artery fusion. C1 origin of PICA reflects dominance of the C1 radiculopial artery, which via the C1 segment of the lateral medullary artery, gives rise to the PICA. The AICA-PICA balance in extent of cerebellar territory capture is a consequence of either anterior spinal (AICA) or lateral spinal (PICA) dominance. All of these cases are given angiographic illustrations below.
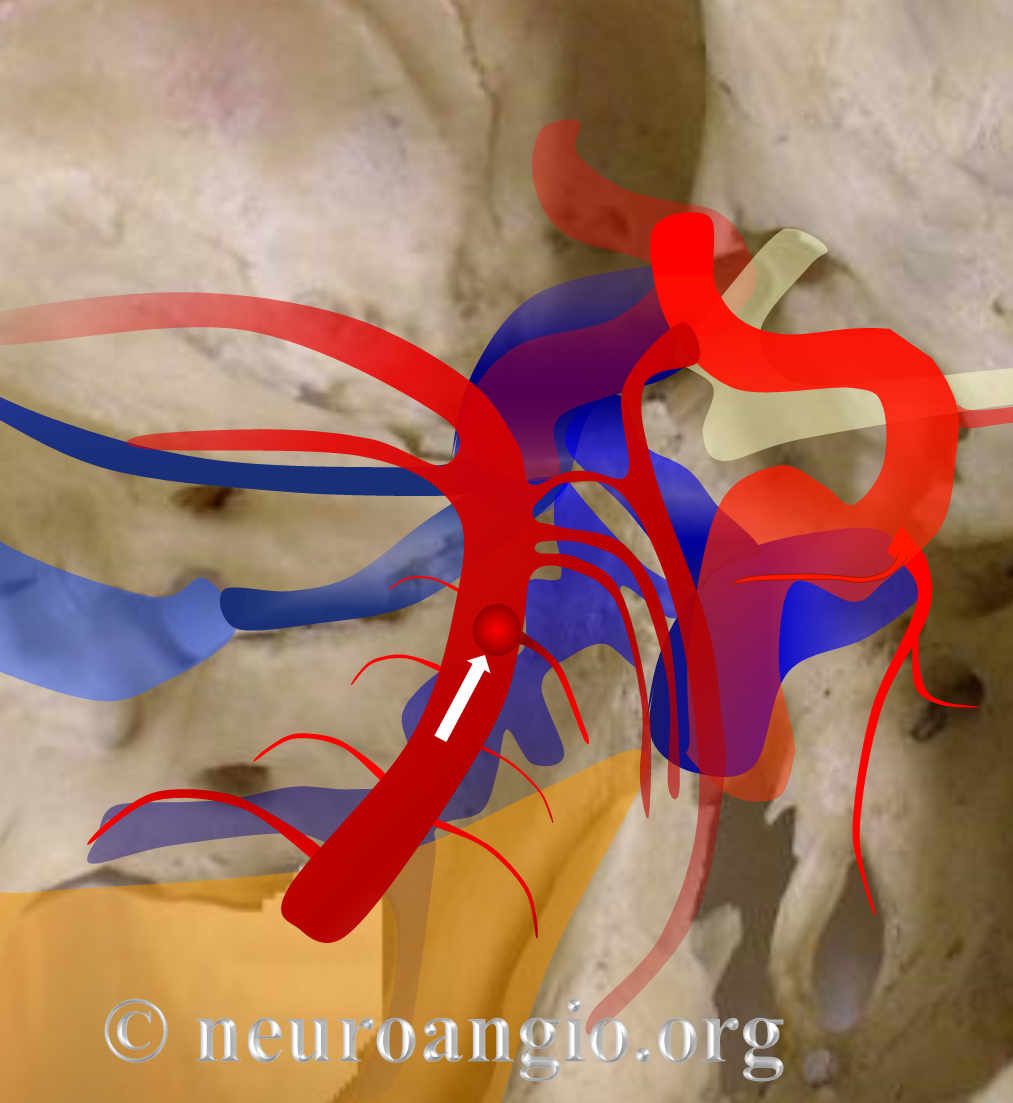
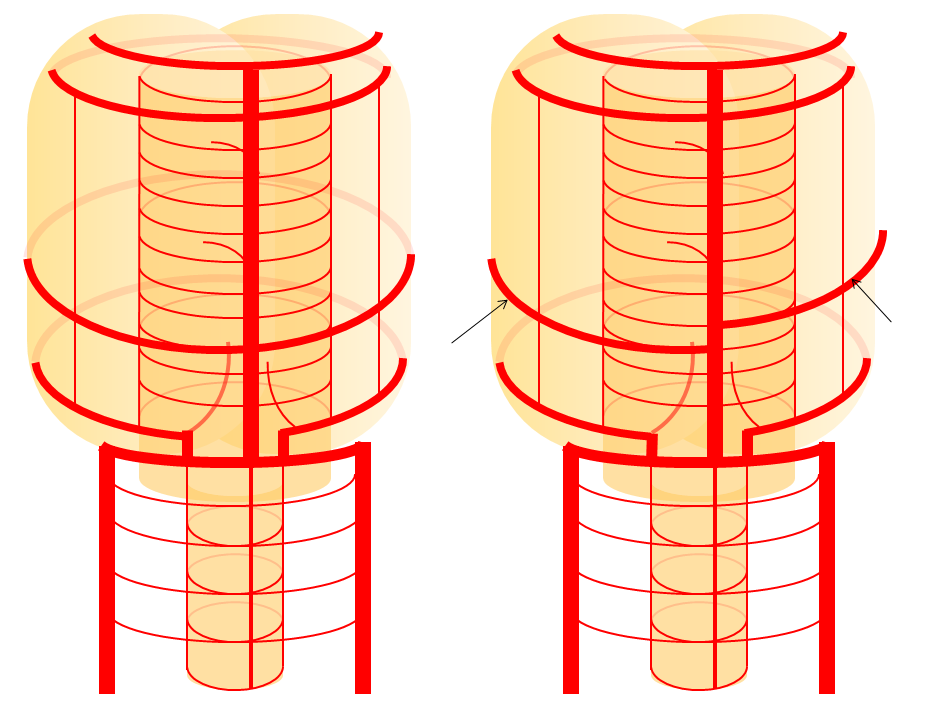
Basilar artery perforators Classical dispostion depicts AICA and SCA arising from the basilar artery, in addition to multiple short basilar perforators whose supply is limited to the brainstem. Understanding the embryology of vertebrobasilar circulation helps explain many variations seen in this pattern. As explained above, one can think of the basilar as a longitudinal channel, with multiple trasverse channels. As the cerebellum develops, some of these transverse channels enlarge to capture the cortical territory of the cerebellum — thus becoming AICAs and SCAs. Others stay relatively small and are thus “limited” to brainstem supply. This simple concept explains ALL variations seen in the area. For example, duplicated SCAs and AICAs are just two adjacent perforators, which persisted in co-dominance of cerebellar supply. Dominant AICAs and PICAs and SCAs are simply variations in extent of cerebellar surface capture by one vessel, with corresponding dominance or hypoplasia of the others. “Larger” vs. smaller basilar perforators are but transverse channels which may capture some small segment of anterior cerebellar surface that is not taken by adjacent AICAs or SCAs . Also important to recognize is the fact that brainstem perforators usually “arise” from SCA and AICA — or, rather, that AICA and SCA in fact “arise” from such perforators. Important, though very small and thus typically angiographically invisible, collaterals exist between these perforators. It is probable that extent of such collateral supply underlies the tremendous clinical recovery occasionally observed following ischemic brainstem infarcts. Almost always, one or more “large” perforator is present between the AICA and the SCA — so consistently visualized, in fact, that it might almost deserve a unique name (we thought to name it “ziggy” — or rather “Arteria ziggus basilarius” — until we found out that it already has a name — “Transverse Pontine Artery”). Occlusion of these transverse pontine arteries typically corresonds to ventral pontine infarction, and may lead to the ischemic cause of dreaded “locked in” syndrome. Since these vessels are in balance with the AICA, dominant PICA with relative AICA hypoplasia is usually accompanied by relative prominence of these perforators (red oval).
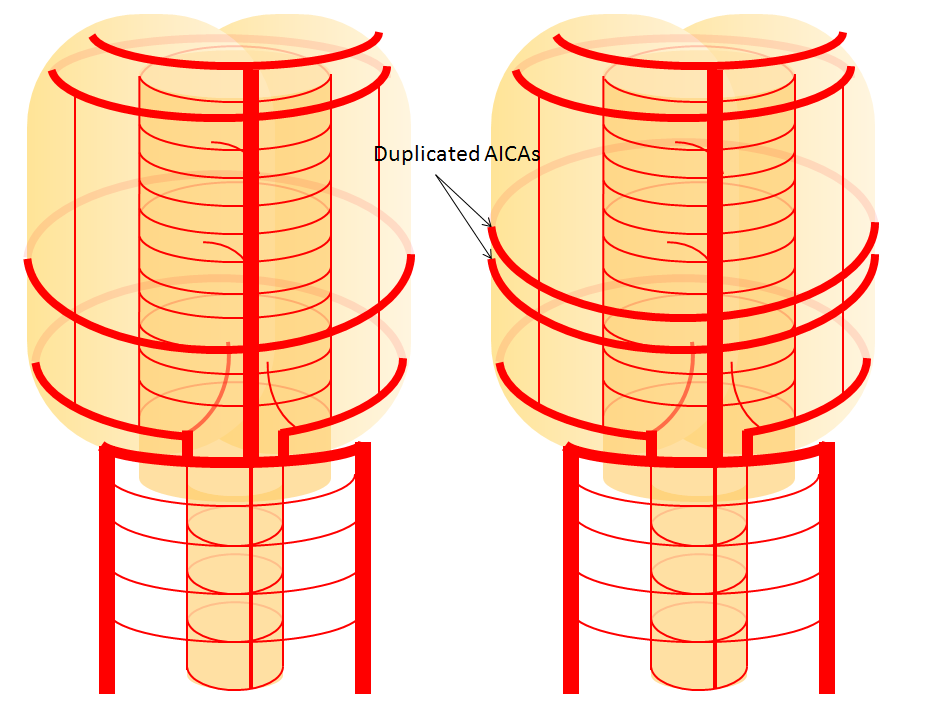
Diagram of the above concept. Image on left, with brainstem alone, shows schematic of vertebrobasilar system with numerous “transverse” perforators supplying the brainstem (transverse pontine arteries). Image on right, with cerebellum in place, depicts SCA, AICA, and PICA as perforators which elnarged to capture cerebellar hemispheric territory. This viewpoint allows for ready conceptualization of multiple SCA, AICA, and PICA variations observed within the vertebrobasilar system.
Perforators and “Nonaneurysmal” but Really Aneurysmal Perimesencephalic Hemorrhage
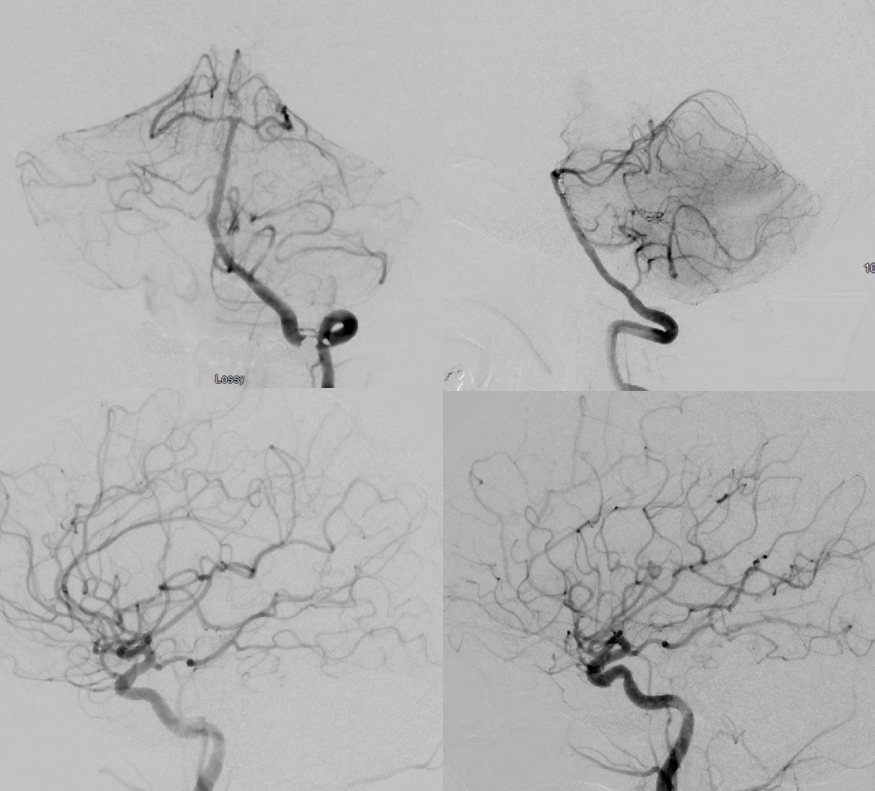
There is ample evidence, much of it gathered at Bellevue and NYU by our group, that many “nonaneurysmal” perimesencephalic hemorrhages are caused by basilar perforator aneurysms. These are likely dissecting. Because the perforators are tiny, the hemorrhages tend to be limited. Prognosis is excellent — as it is for most perimesencephalic bleeds — that we knew. The difference is now we know how many of them bleed. See links to specific cases here 123
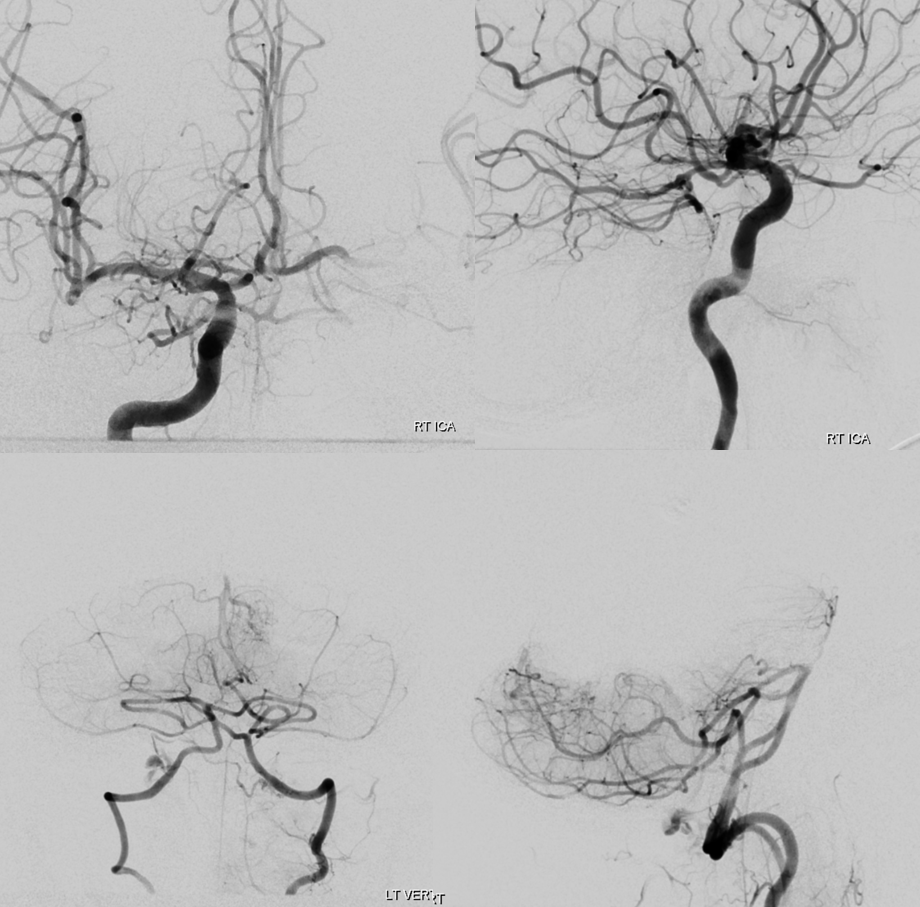
Below are examples
Examples
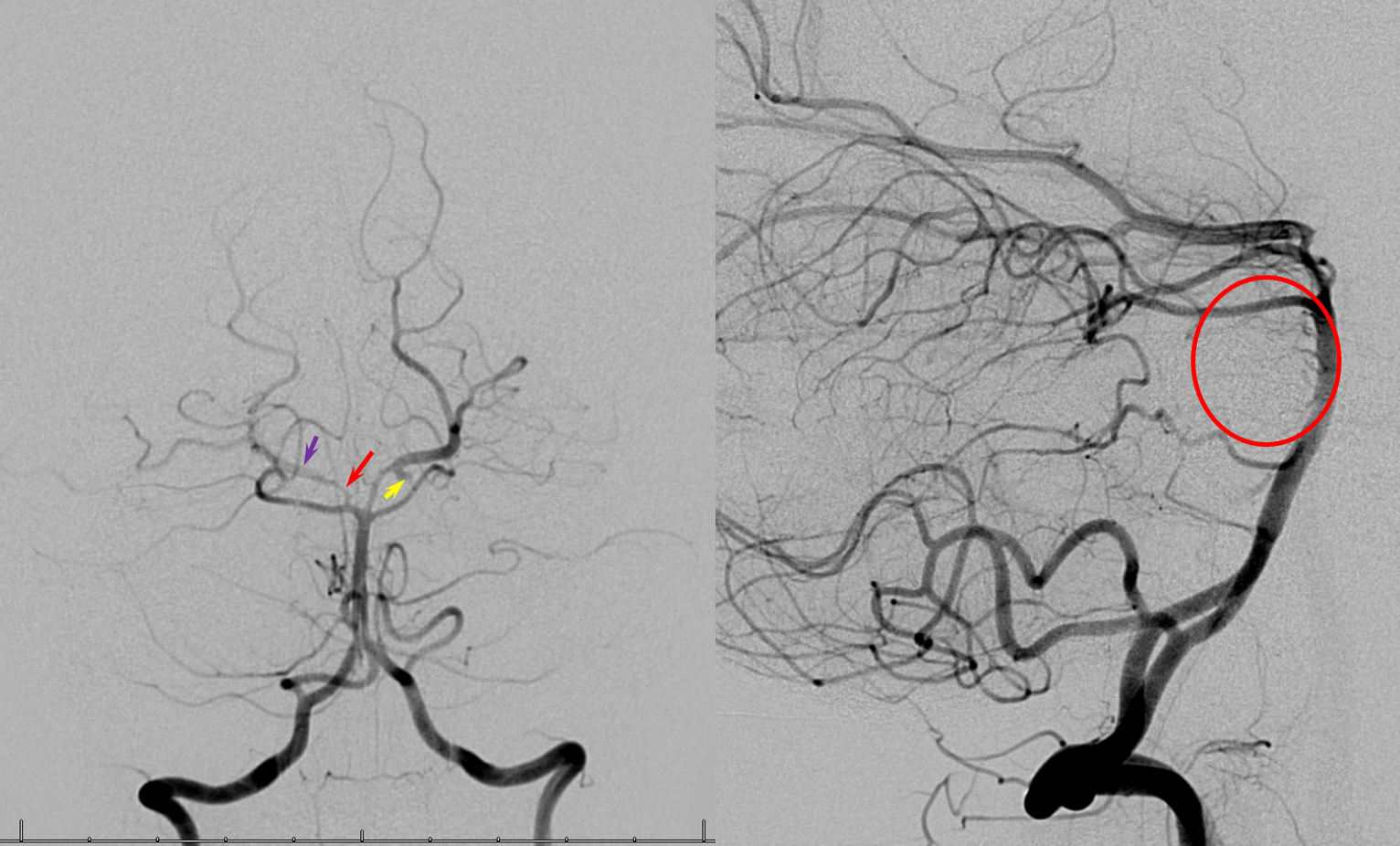
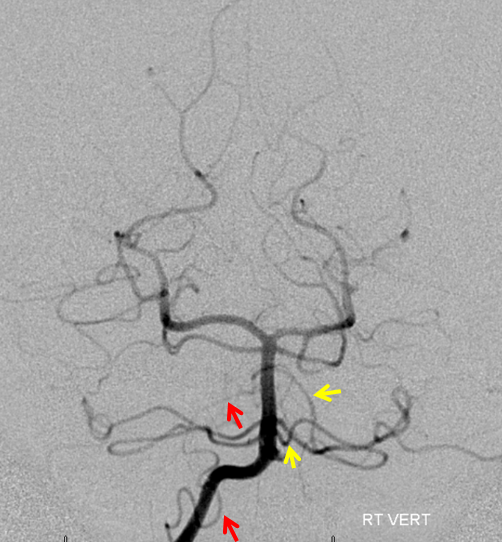
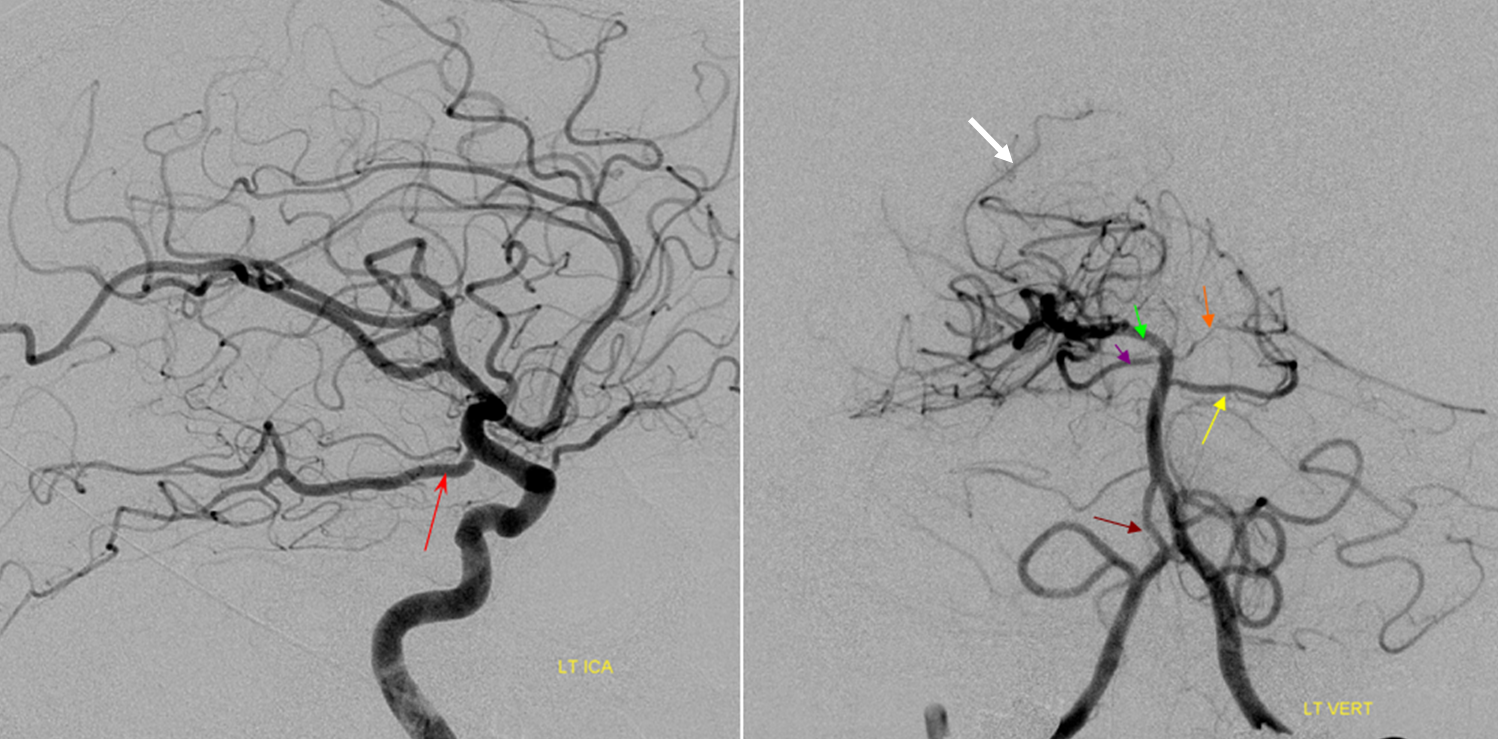
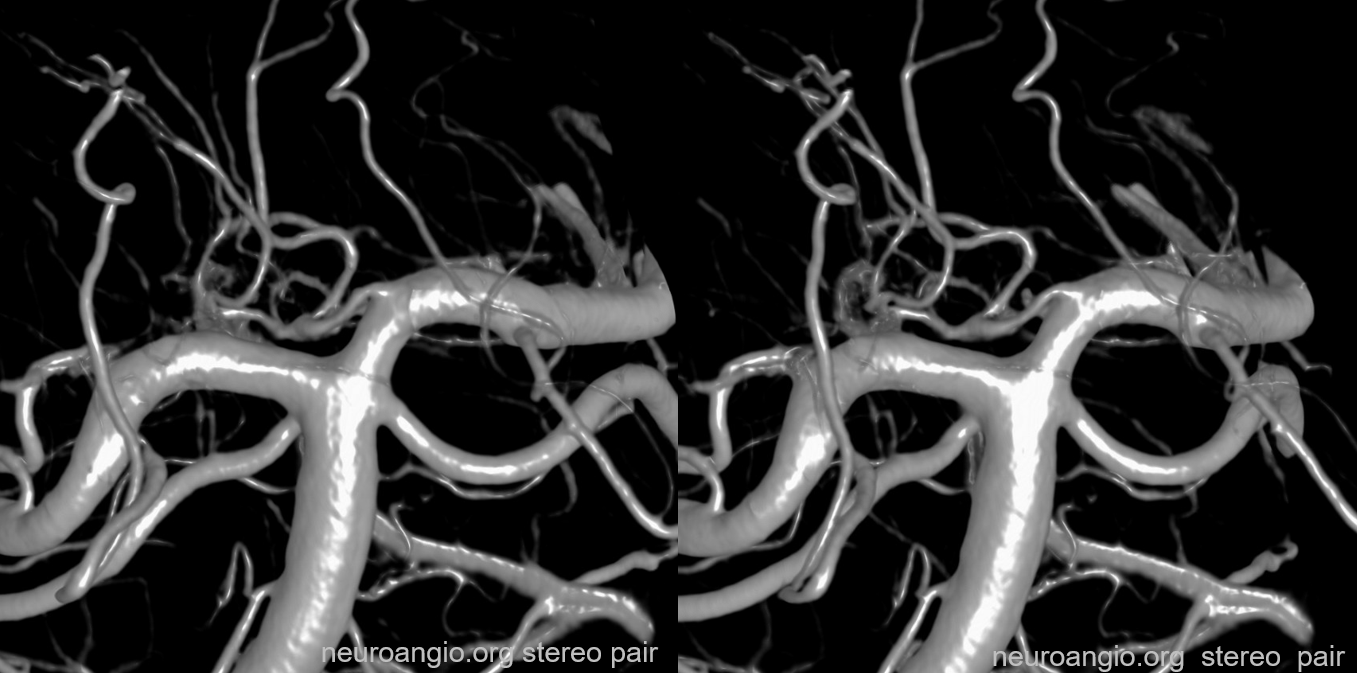
Duplicated AICAs — co-dominance of two transverse pontine arteries in supply of cerebellar territory.
Schematic of the same disposition. AICA-PICA balanceAn exceedingly well known variation — a continuum, in fact, where AICA and PICA are, respectively, larger and smaller, depending on the extent to which one can capture the territory of the other. The PICA can be conceptualized as a large radiculopial origin vessel (arising from the lateral spinal artery, a homolog of the posterior spinal system), whereas AICA originates as coronary type vessel off the basilar (homolog of anterior spinal artery).
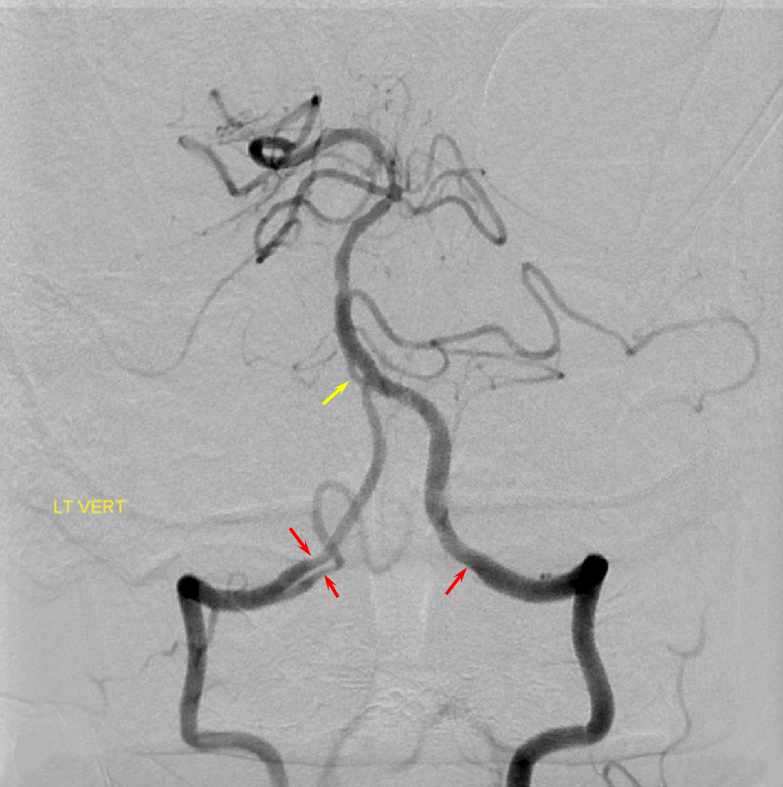
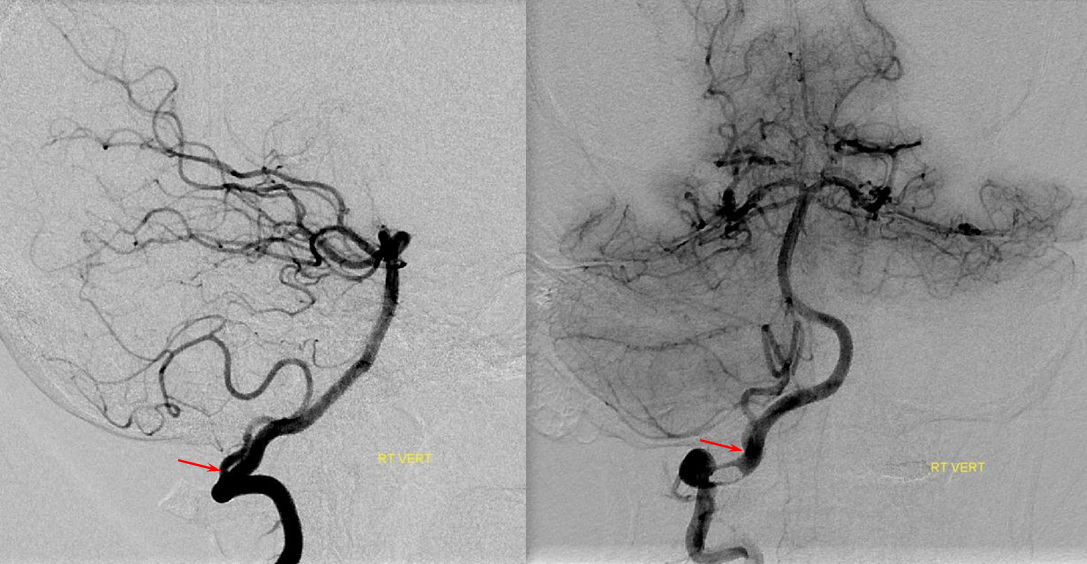
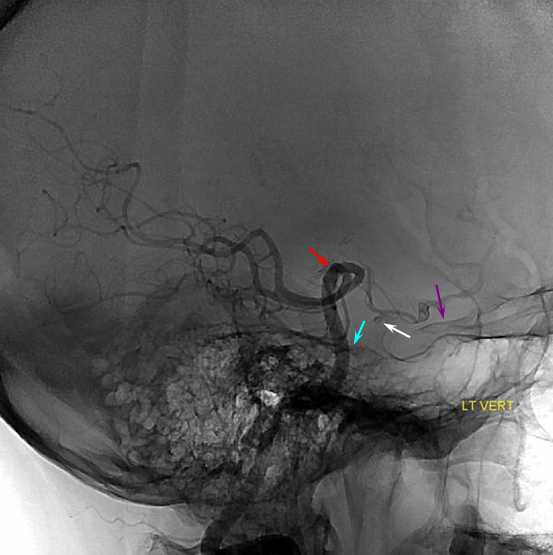
Schematic of balanced arrangement (left image). Right image shows PICA dominance of the “right” vert and AICA dominance of the “left” vert. Below is an angiographic image of the same arrangement, except for a kind of AICA duplication on the left, and a small basilar fenestration (yellow arrow). Notice how PICA and vert change caliber when they go through the dura above foramen magnum. Not to be confused for atherosclerotic disease, of which there is also a small specimen present just above the AICA.
Superior Cerebellar Artery Duplication — same as AICA, just a balance of transverse arteries.
Big picture overall analysis of branching pattern AICA PICA lateral spinal
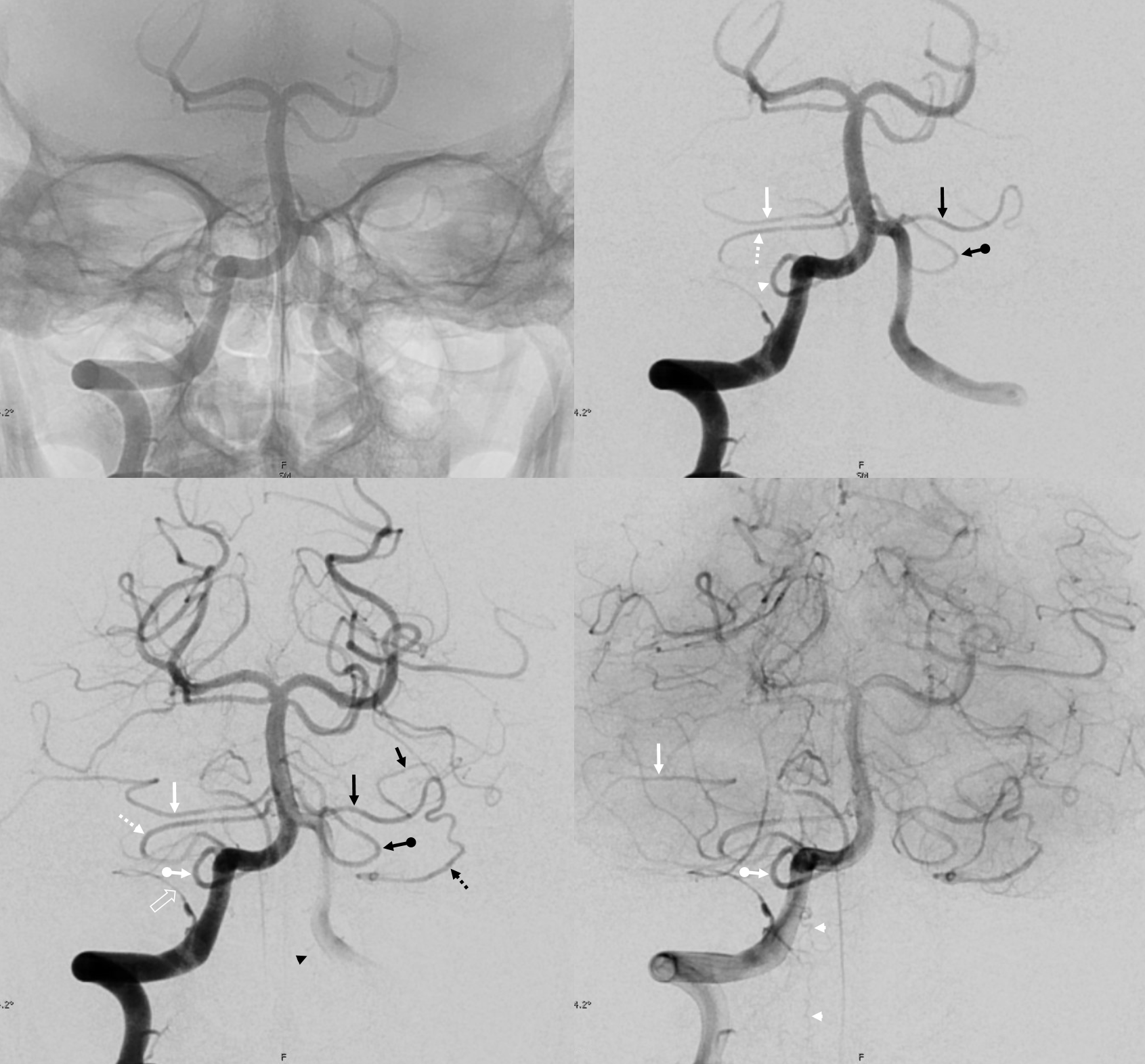
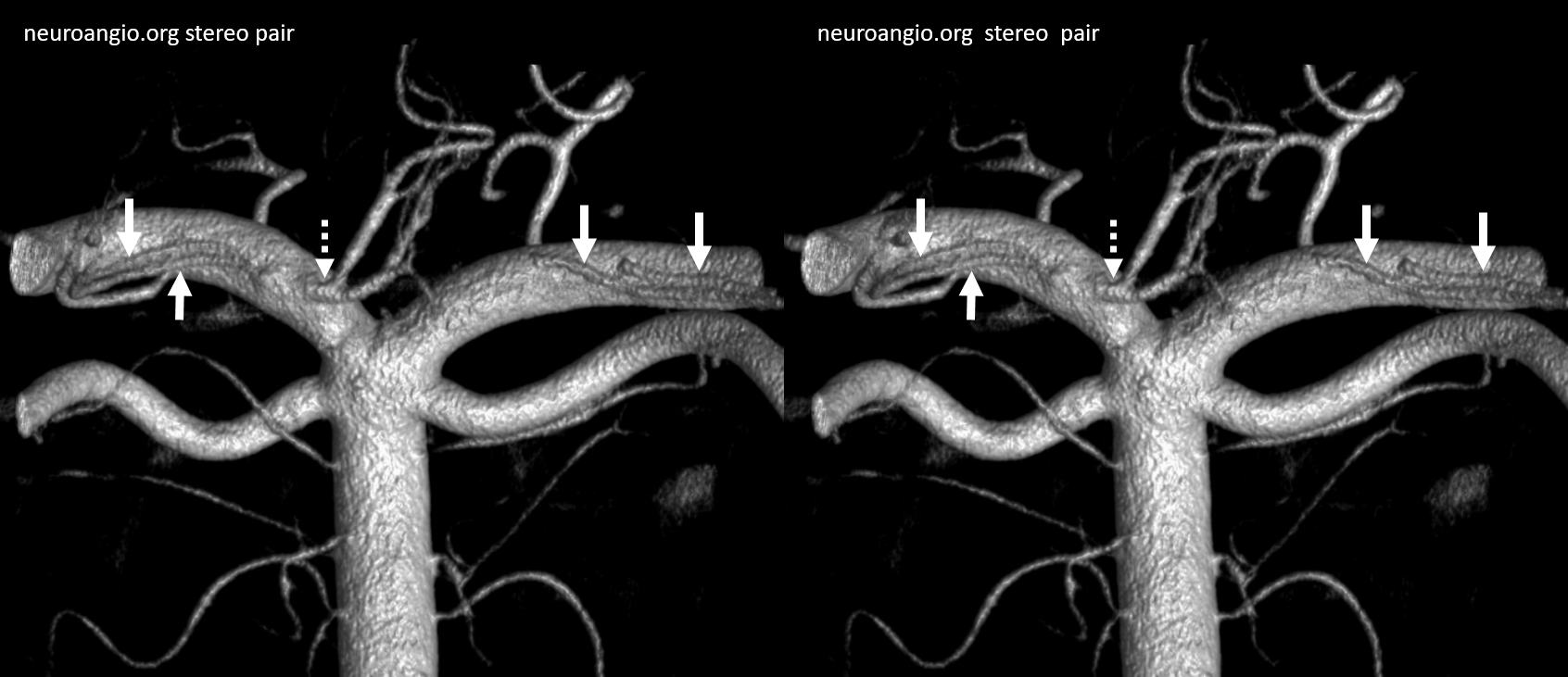
The “balance” approach to anatomy we emphasize requires considering different vascular territories as part of the whole. In the example below, this analysis reads something like this: on the right, there is an apparent “duplicated AICA”. Thats not really true. The AICA, as evidenced by its lateral course towards the IAC area, is the upper vessel (white arrow). The lower vessel (dashed arrow) is a PICA territory artery supplying the inferior cerebellar territory. the lower-most vessel is also a PICA territory artery (ball arrow) supplying the vermian territory. A large lateral spinal artery (arrowhead) is also present, since there is dominant right PICA. There is a posterior meningeal artery also (open arrow) On the left, the dominant AICA (black arrows) supplies both AICA and inferior hemispheric territory of the PICA. The lower vessel is PICA with supply limited to vermian territory. A very small lateral medullary artery (arrowhead) is also present.
Pathoanatomy
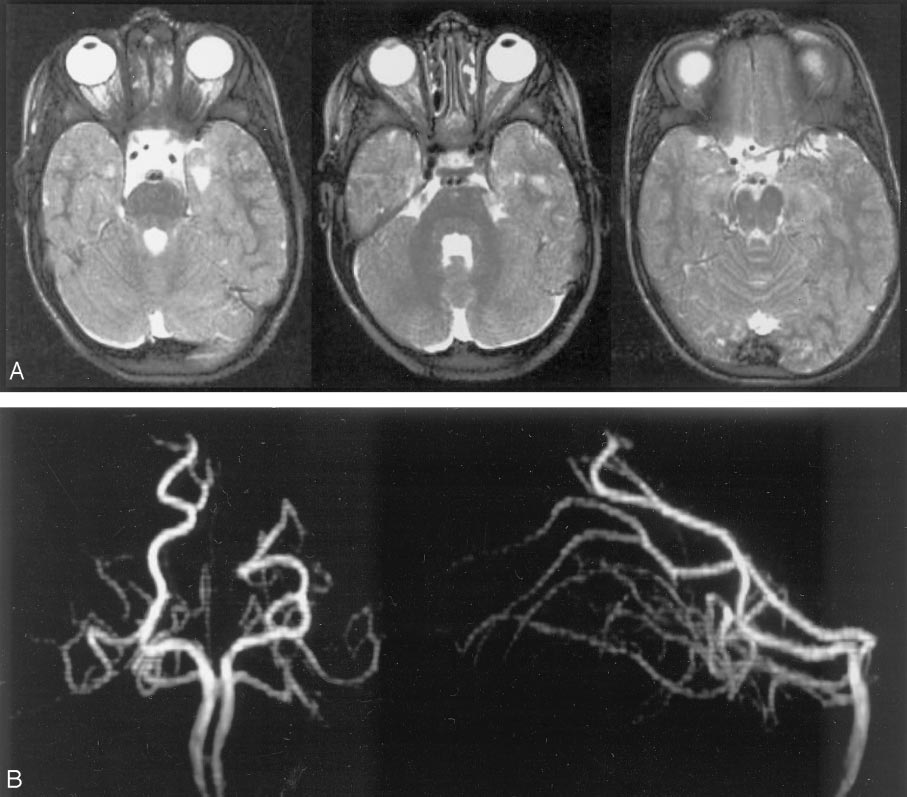
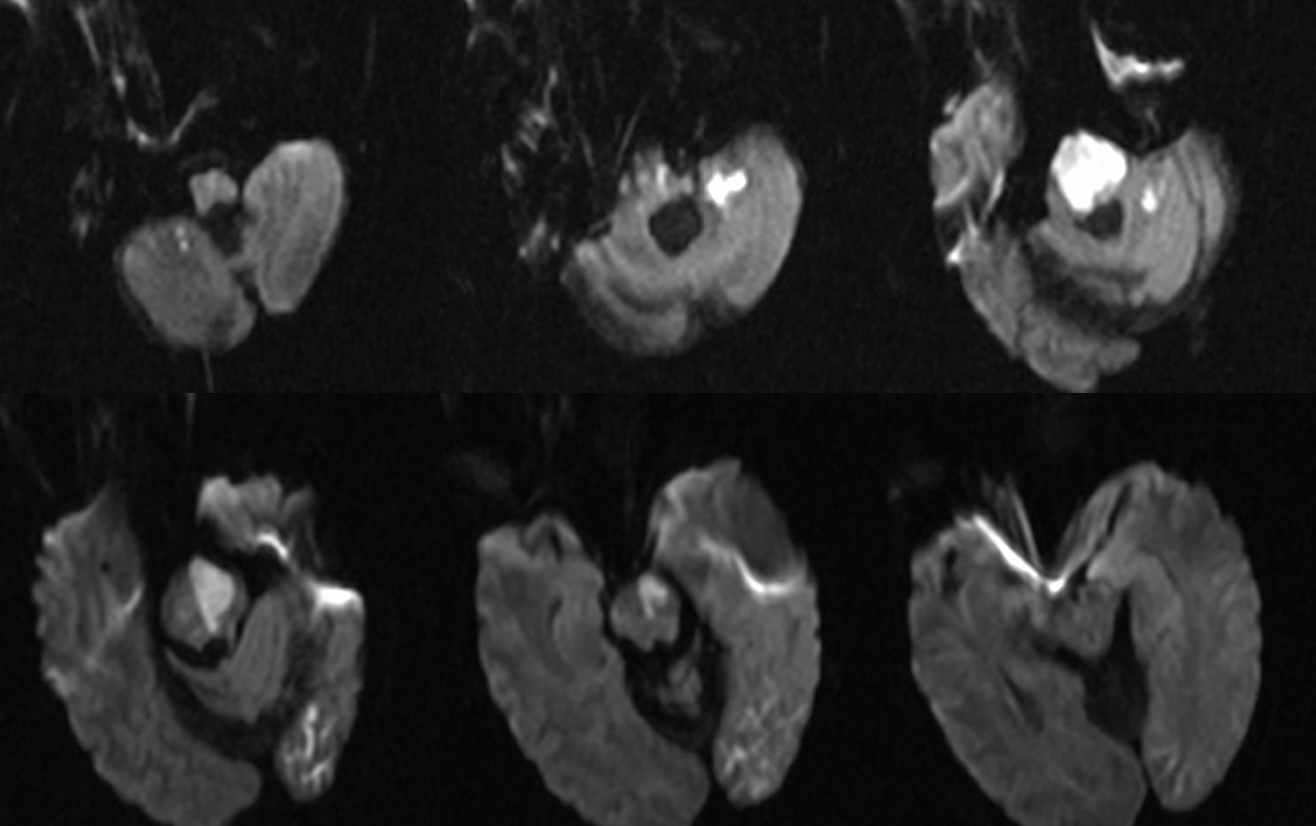
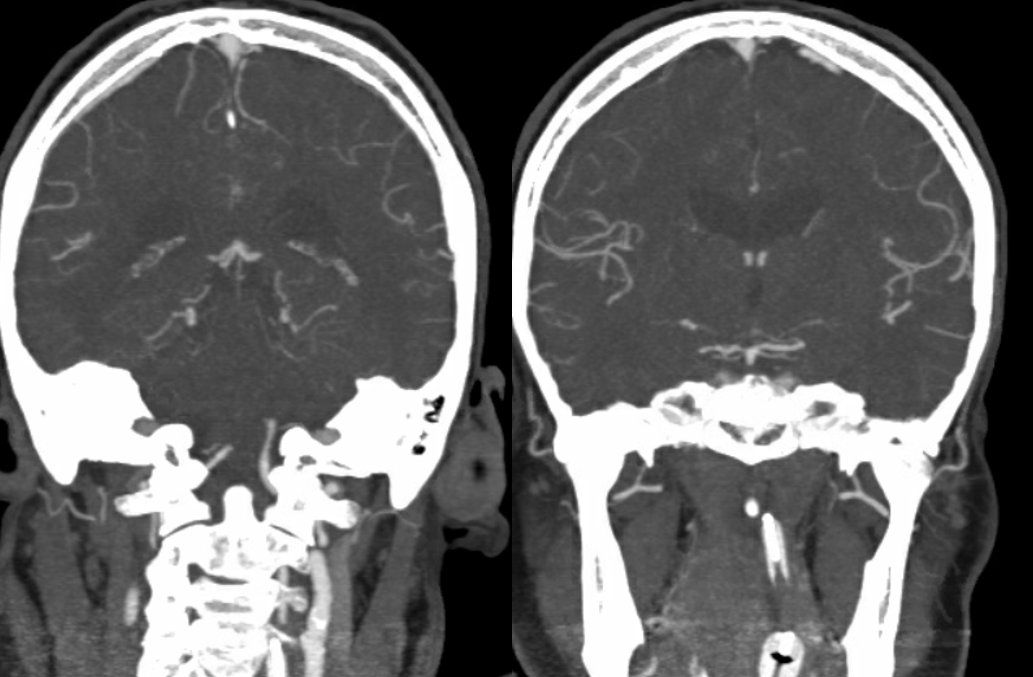
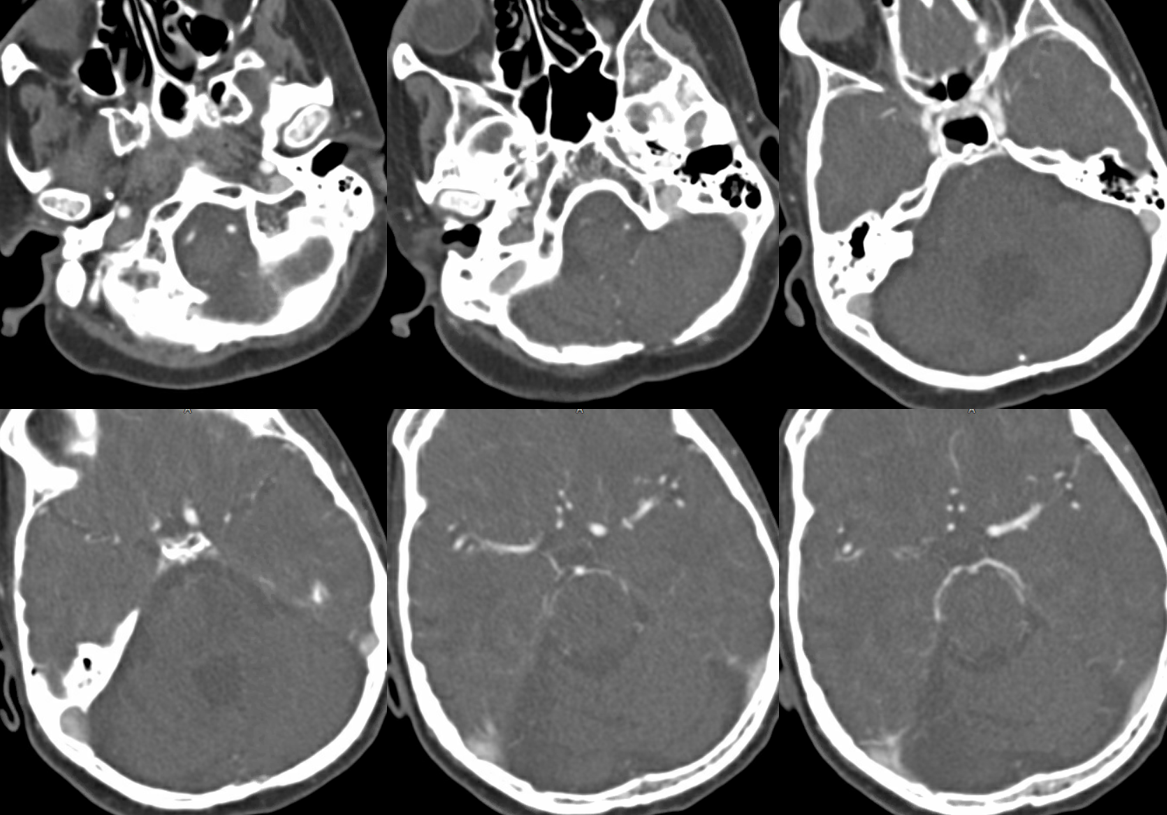
This remarkable case illustrates how properly interpreted pathology offers proof of “theoretical” concepts. A ruptured AVM in the ventrolateral midbrain is supplied by two hypertrophied transverse pontine arteries, rather than AICA or SCA. What an unusual picture! Why? The AVM was present, in microscopic form, in utero. Flow demand by the shunt was met by twin transverse spinal arteries, which remained the principal supply of the AVM, while AICA and SCA developed to supply normal brain territory, with minor contributions to the AVM. Interestingly, both arteries were embolized to “stumps” off the basilar artery, with no neurologic deficit. Noncontrast head CT with blood in the lateral ventricles (yellow), and CTA demonstrating nidus (red) and some associated aneurysms (purple)
Angiographic views of vertebrobasilar circulation, showing two hypertrophied transverse pontine arteries (white arrows) supplying an AVM, with associated perinidal arterial aneurysms (purple). Image on left is pre-embo, center is status-post embo of the more cranial transverse pontine pedicle, image on right is post-embo. Also shown are the SCA (pink) and AICA (black). Notice an area of spasm involving the more caudal transverse pontine artery (blue arrow) center image (the more superior transverse pontine feeder has been glued).
Post-embolization CT:
A schematic of this situation is shown below:
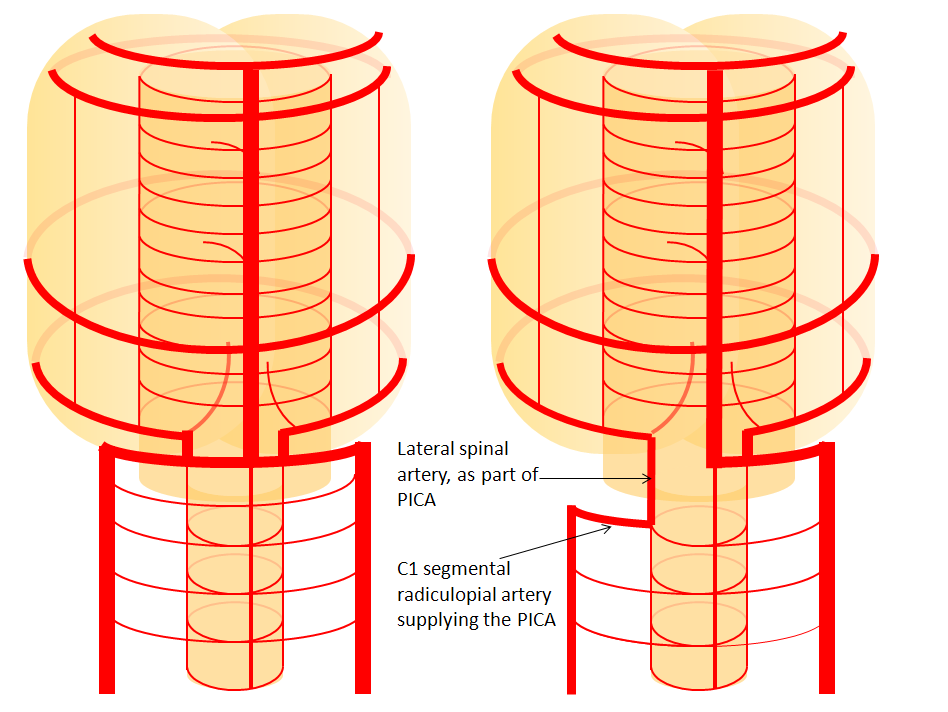
C1 origin PICA — since PICA can be regarded as a branch of the lateral spinal system, its origin may vary from usual to lower down along the lateral spinal line — C1, for example. In this case, the vertical segment of the PICA, extending from C1 through the foramen magnum, corresponds to the “lateral spinal” part of PICA.
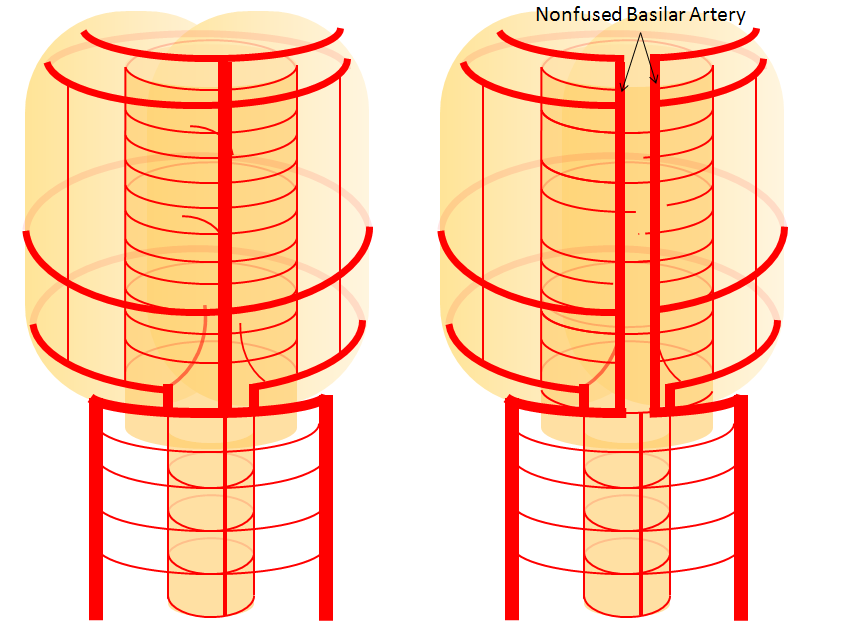
Variation in fusion patterns of the basilar artery
As mentioned above, the basilar artery is formed via coalescence of multiple channels belonging to the longitudinal neural system, following development of the anterior circulation (see Neurovascular Evolution and Vascular Neuroembryology sections) This process sets the stage for multiple basilar artery variations, which can be considered from the standpoint of 1) extent of fusion 2) position of fusion with respect to the brainstem, and 3) completeness of fusion (i.e. fenestrations).
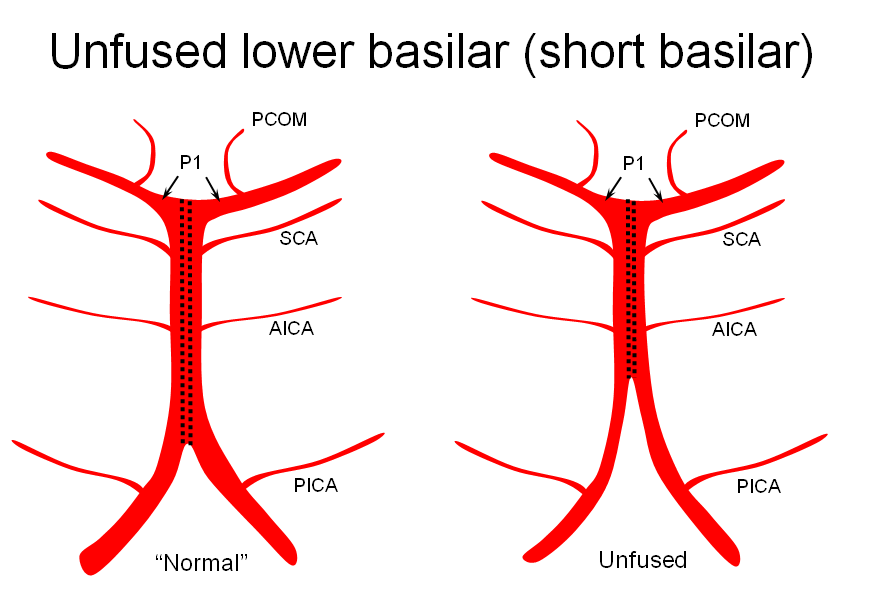
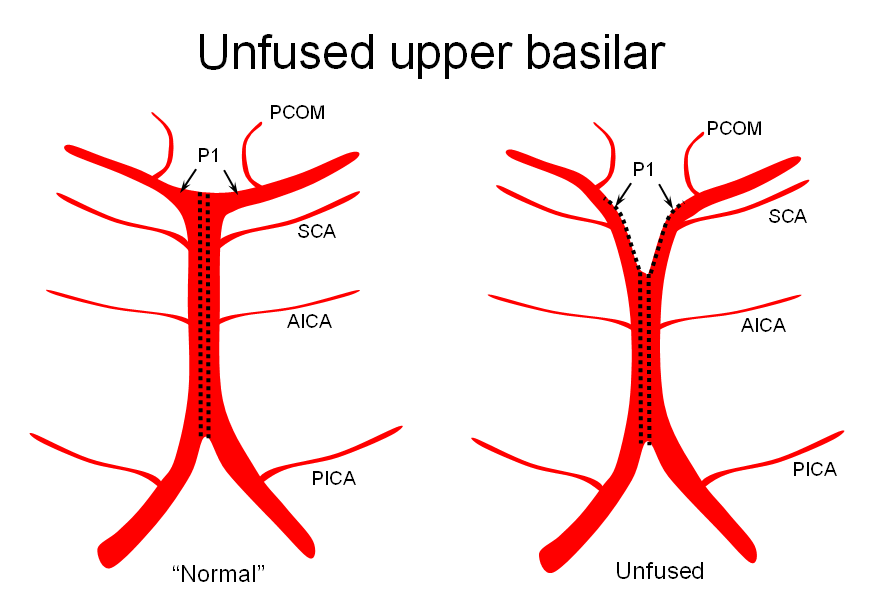
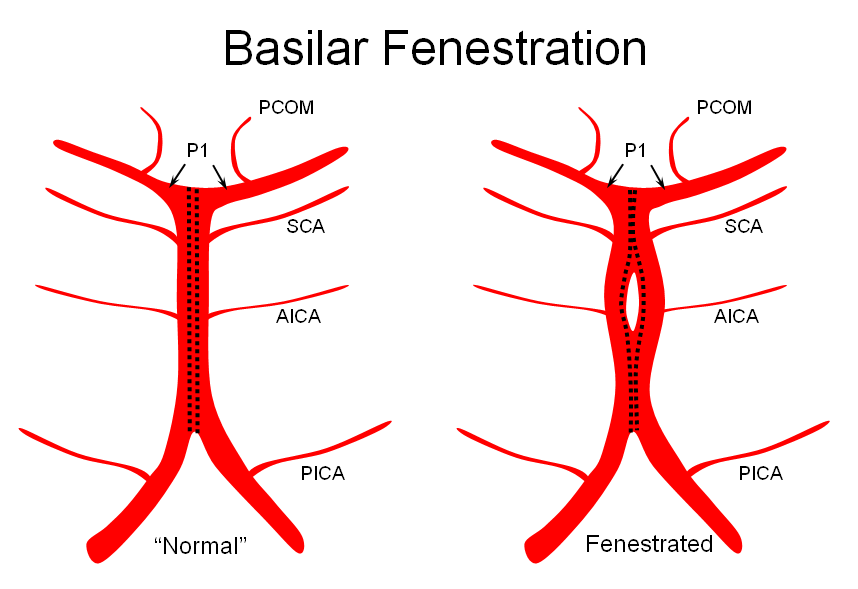
It is conceptually helpful to think of the basilar artery as being “zipped” in the middle, with vertebral and PCA segments being “unzipped”. The length of the “zipped” segment, and the integrity of the zipper determine the final configuration of the artery. As such, the artery may be
1) “short” — corresponding to relative lack of fusion of the caudal or rostral segments
2) Unfused at the top — corresponding to “unzipping” at the basilar tip — less common but much more confusing
3) Fenestrated — broken zipper in the middle
The first one, short basilar, is in fact a continuum in terms of fusion extent. There is no “number” to guide what short or long is, but some basilars are clearly quite foreshortened. (The tortuous basilar of the old hypertensive is a different matter from developmental variation)
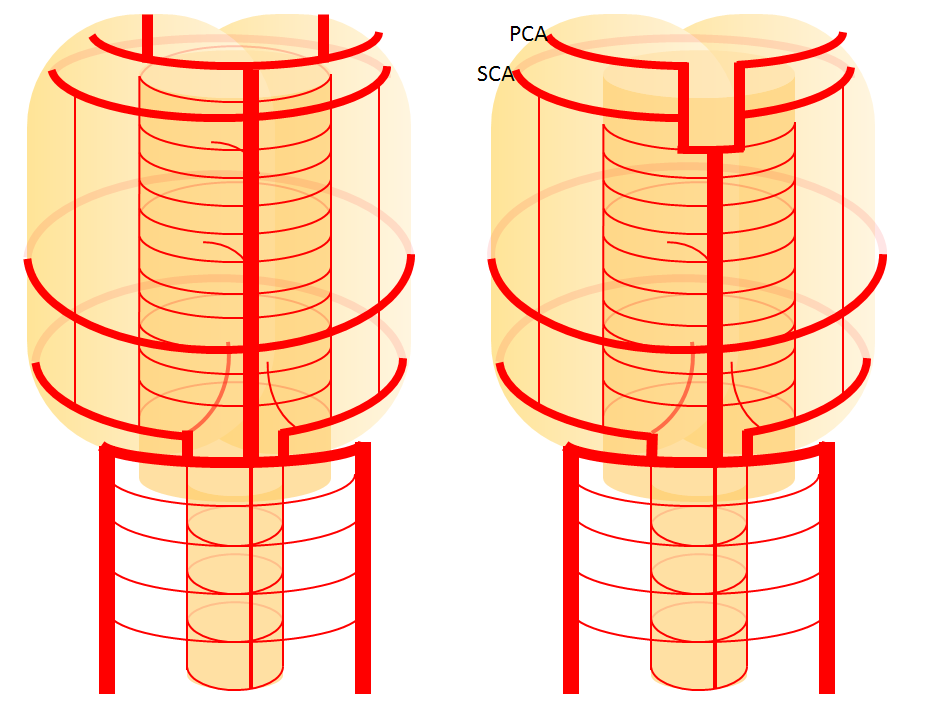
The second variation, involving lack of basilar tip fusion, can generate a lot of confusion. Effectively, the top of the basilar is split in two, so that one or both superior cerebellar arteries originate from the P1 segment. This variant is not, therefore, a primary superior cerebellar artery aberration, but instead a deficiency in basilar fusion.
The third, basilar fenestration, is quite common, and usually of little clinical signficance, except when it is so short as to minic a dissection.
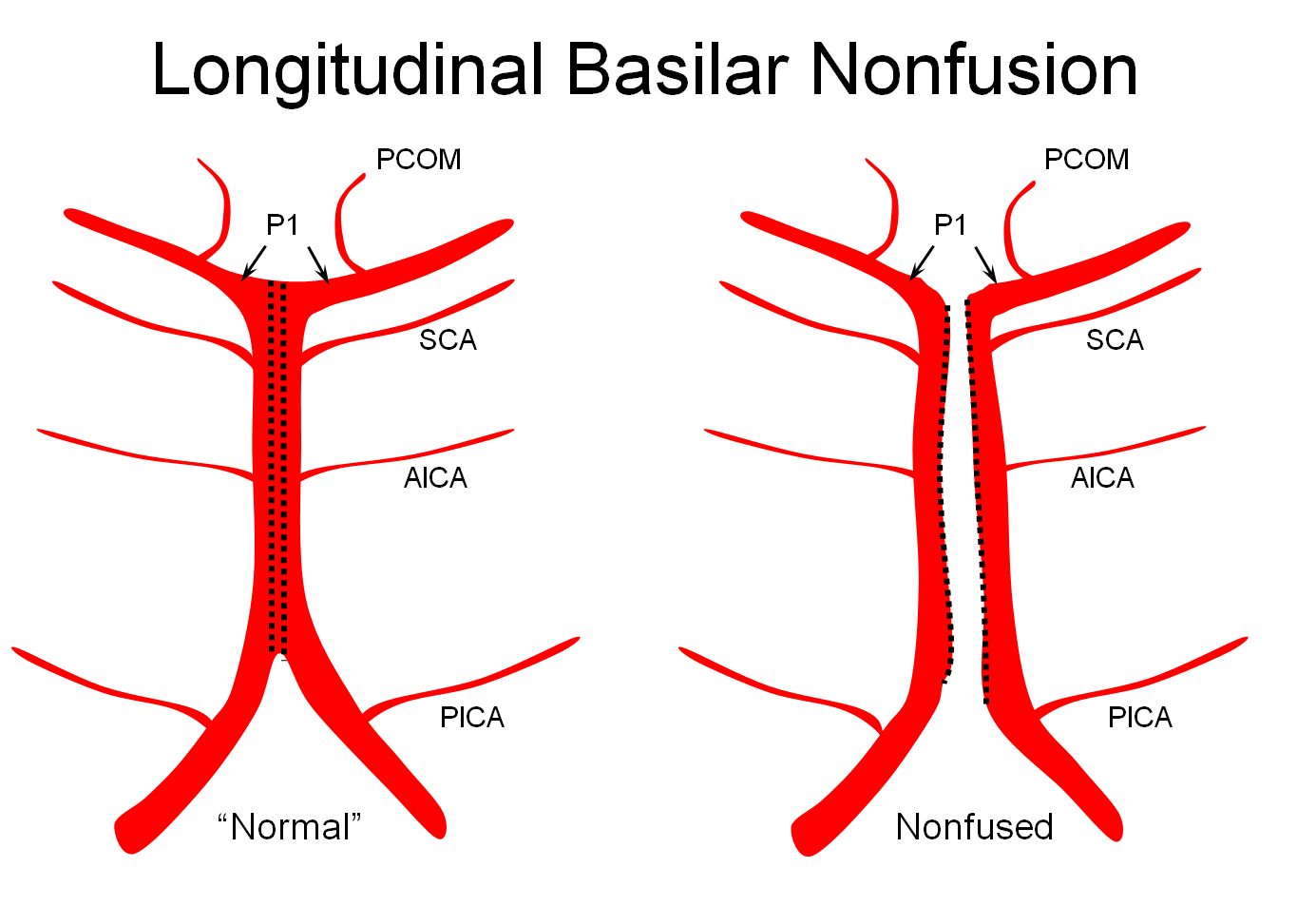
Basilar nonfusion — extreme of fenestration, a completely “unzipped” look. Very rare.
Illustrations
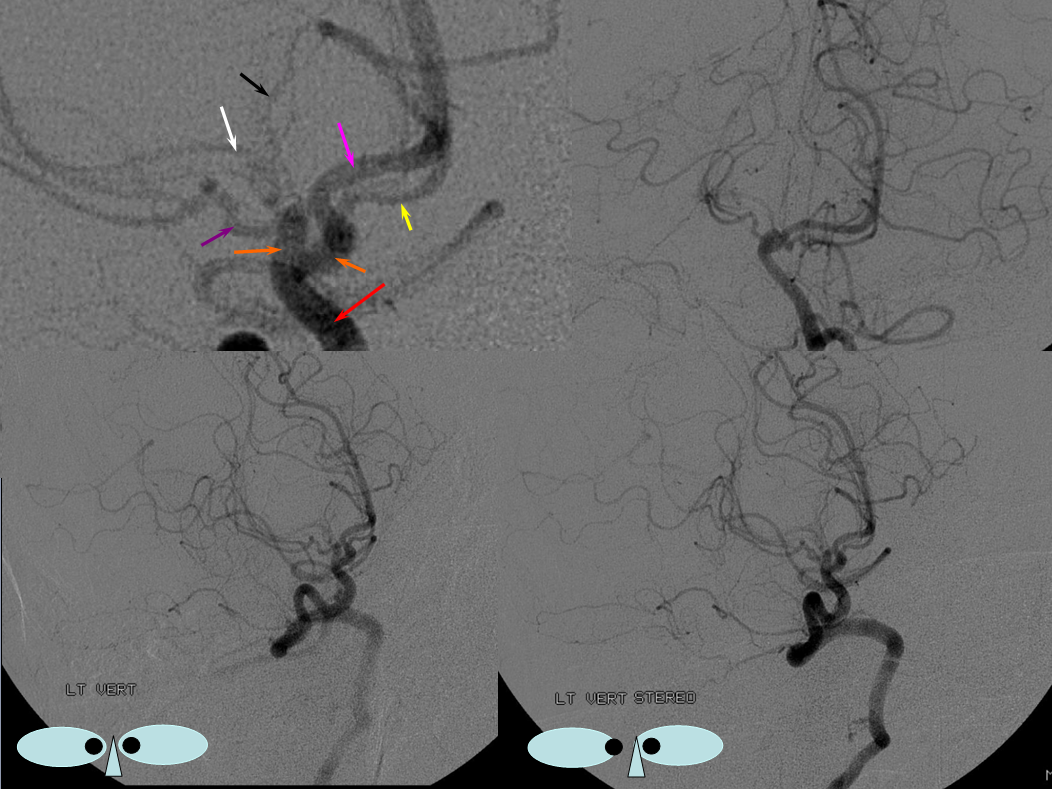
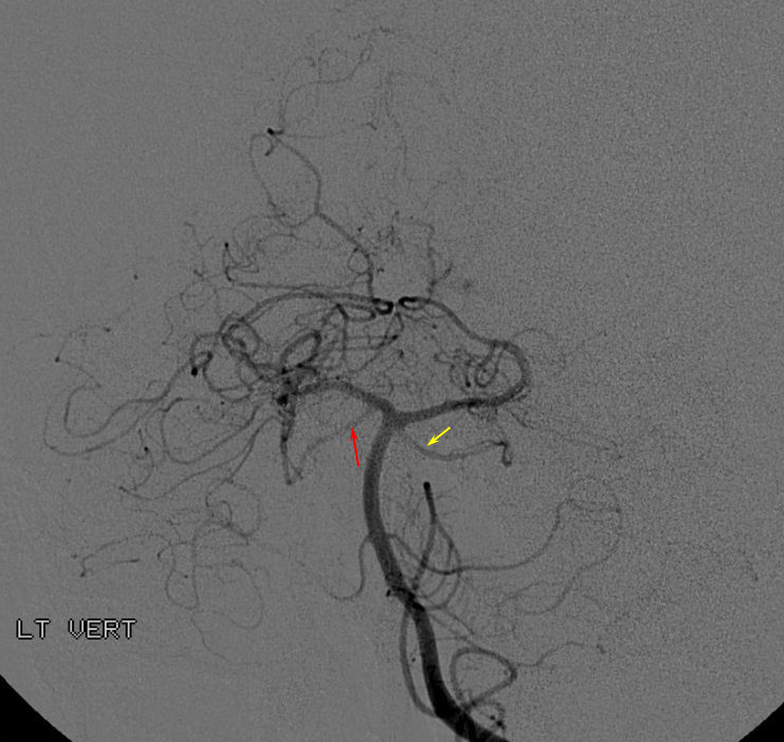
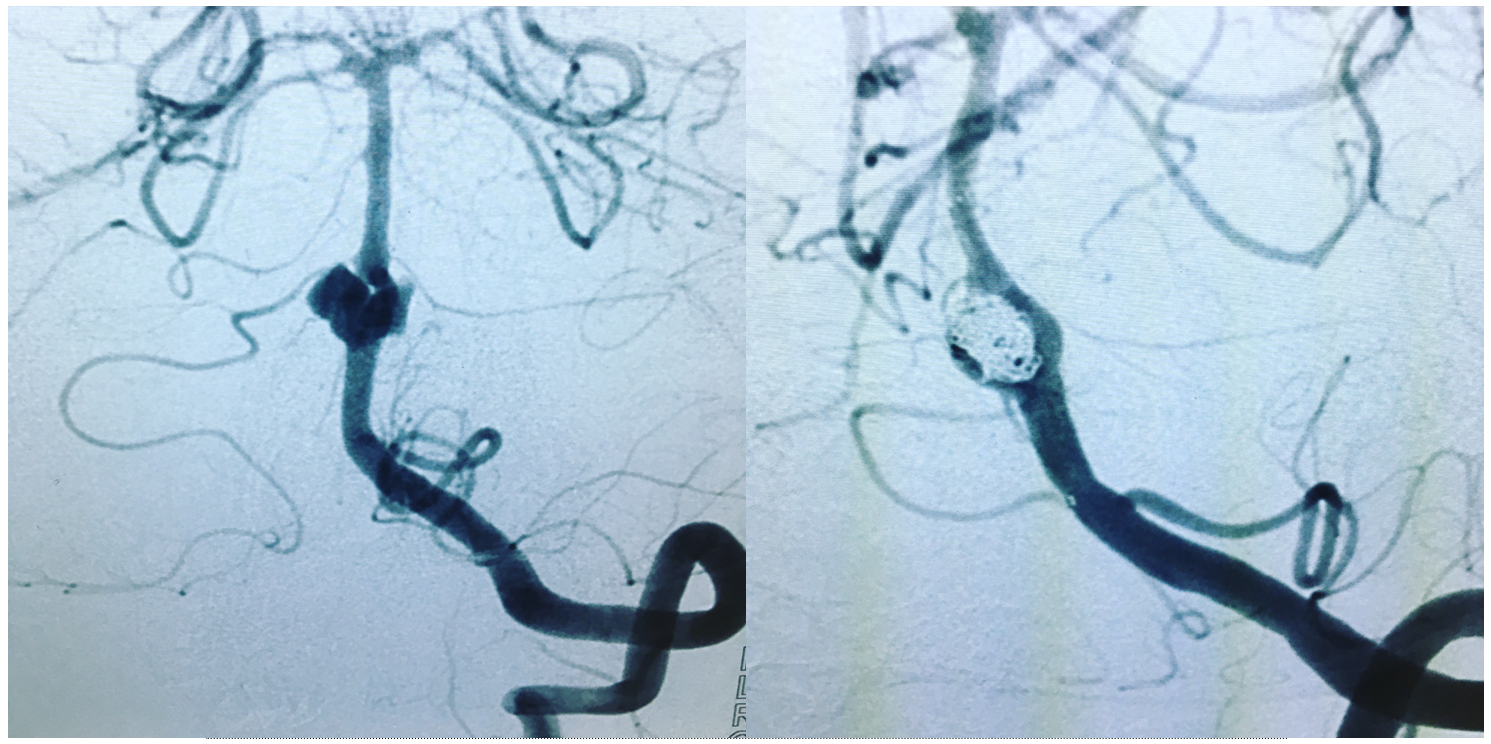
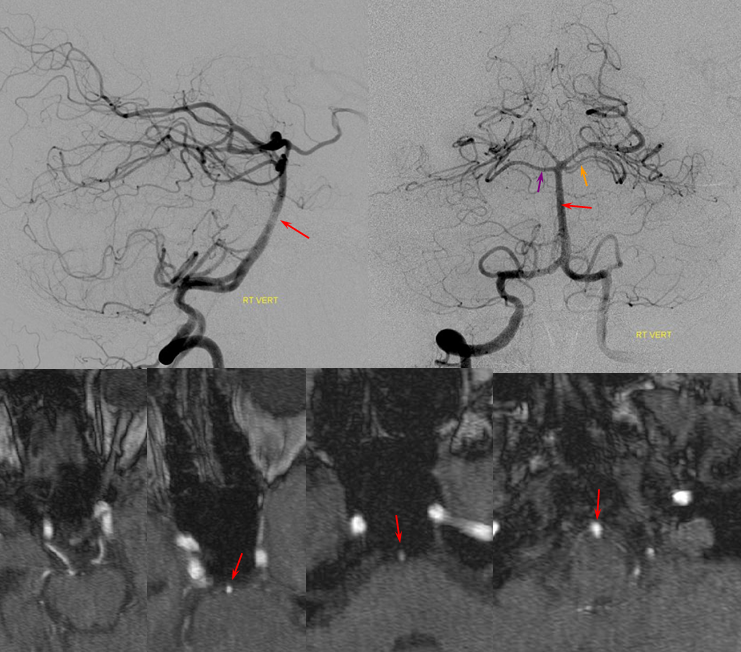
Unfused basilar simulating aneurysm — reported as basilar tip aneurysm on CTA, angio demonstrating an unfused basilar tip with tortuous P1 vessels simulating aneurysm, further complicated by presence of a fetal PCOM on the right.
Origin of superior cerebellar artery at “top” of the basilar on the left (yellow), and from P1 segment on the right (red arrow)– seen often, can be conceptualized as a “short” basilar which did not undergo enough coalescence at the top to incorporate the superior cerebellar artery.
Notice P1 origin of the right superior cerebellar artery (red) with contralateral classical disposition in yellow
The same unfused upper basilar, with the seemingly opposite appearance of the right P1 (red arrow) originating from the SCA. Both cases are in fact variants of deficient upper basilar fusion. P2 wash-in (purple) is present, as well as some reflux into a dominant left PCOM (yellow). Notice abundant pontine perforators (within red oval) in setting of bilateral PICA dominance. The smaller the AICAs, the more perforators will be.
Diagram of P1 origin SCA, as related to deficient fusion of the upper basilar.
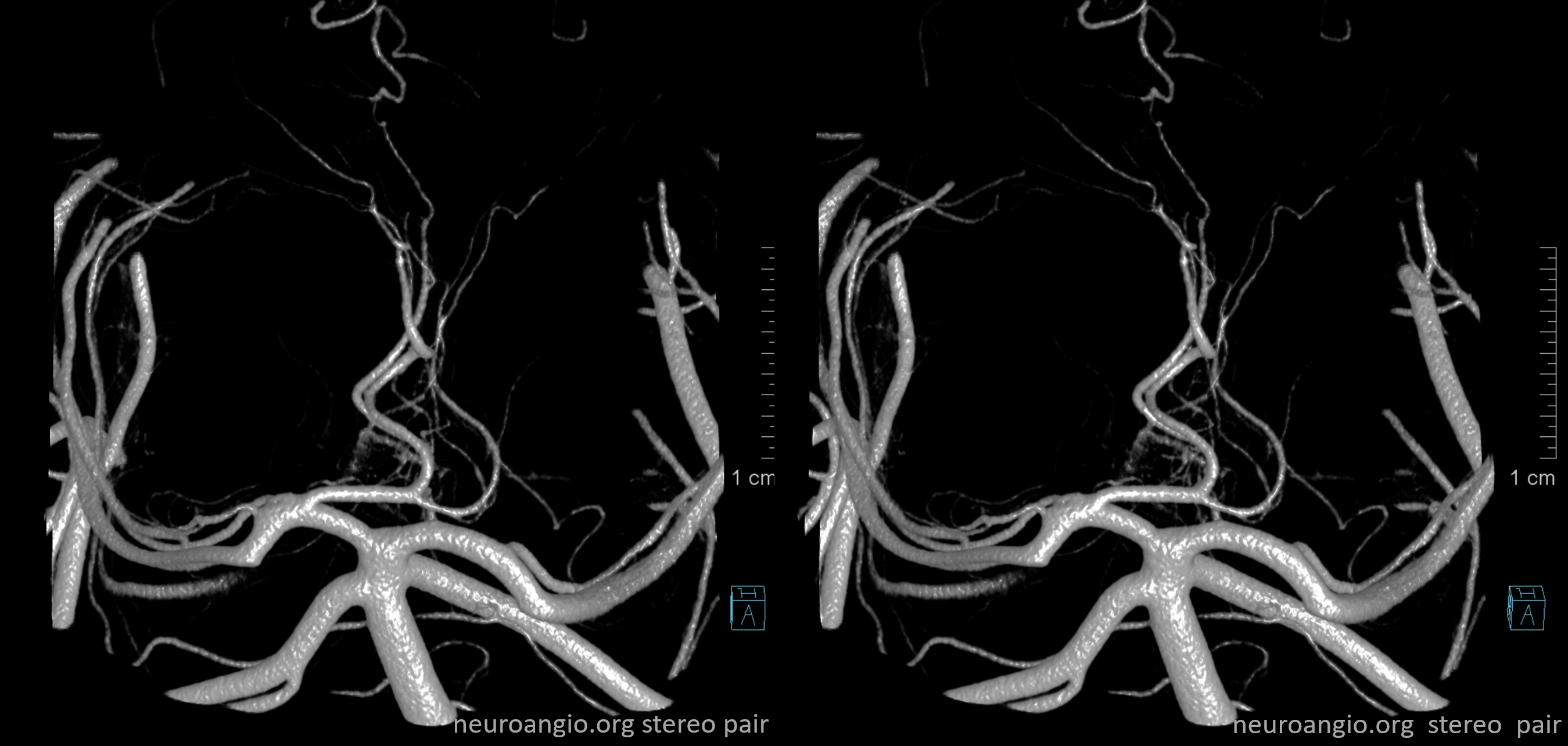
Another illustration, on opposite sides, in a patient with a complete circle of Willis, displayed to advantage by a generous injection.
“Normal” P1 segment origin from distal basilar (in a patient with duplicated AICAs)
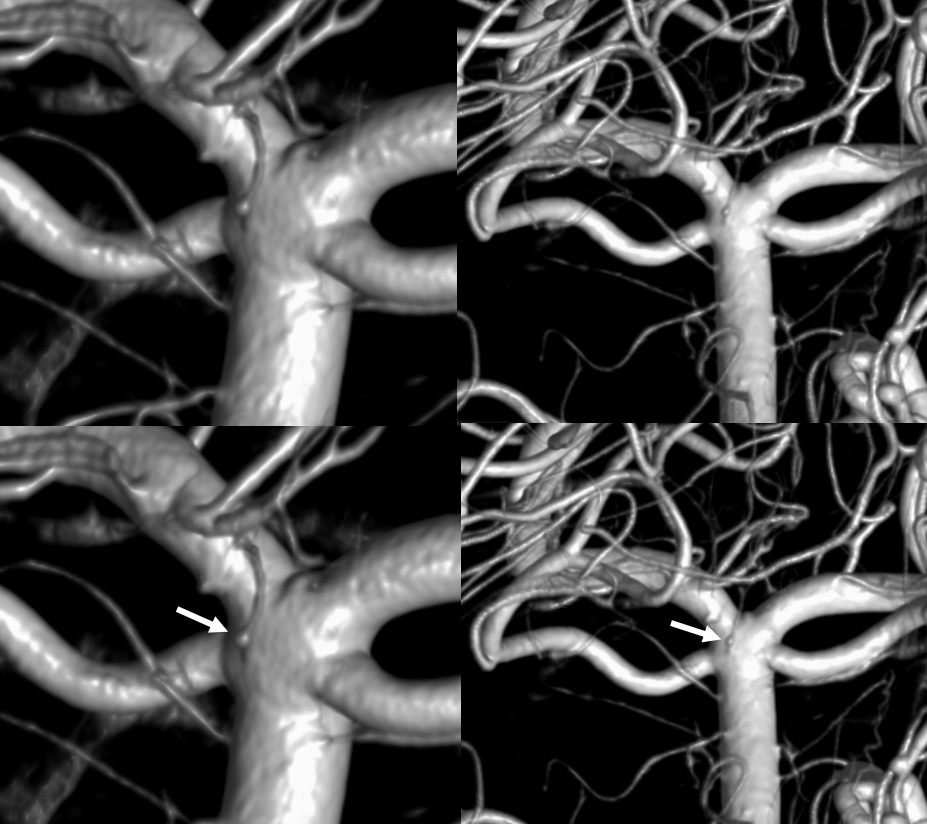
Basilar Artery Fenstration — “broken zipper in middle”. Seen quite often, of litte clinical significance, but illusrative of the fusion concept.
Another fenestration, on CTA
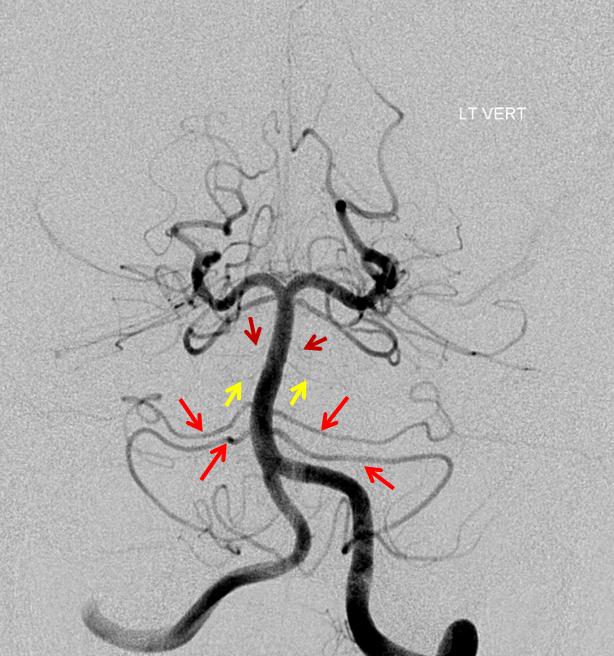
Again, a rather large fenestration (brown arrow), in a patient with fetal PCOM. Notice, how on the right, a small AICA (with dominant ipsilateral PICA) is accompanied by a number of basilar perforators between the AICA and SCA (not marked with any arrows)
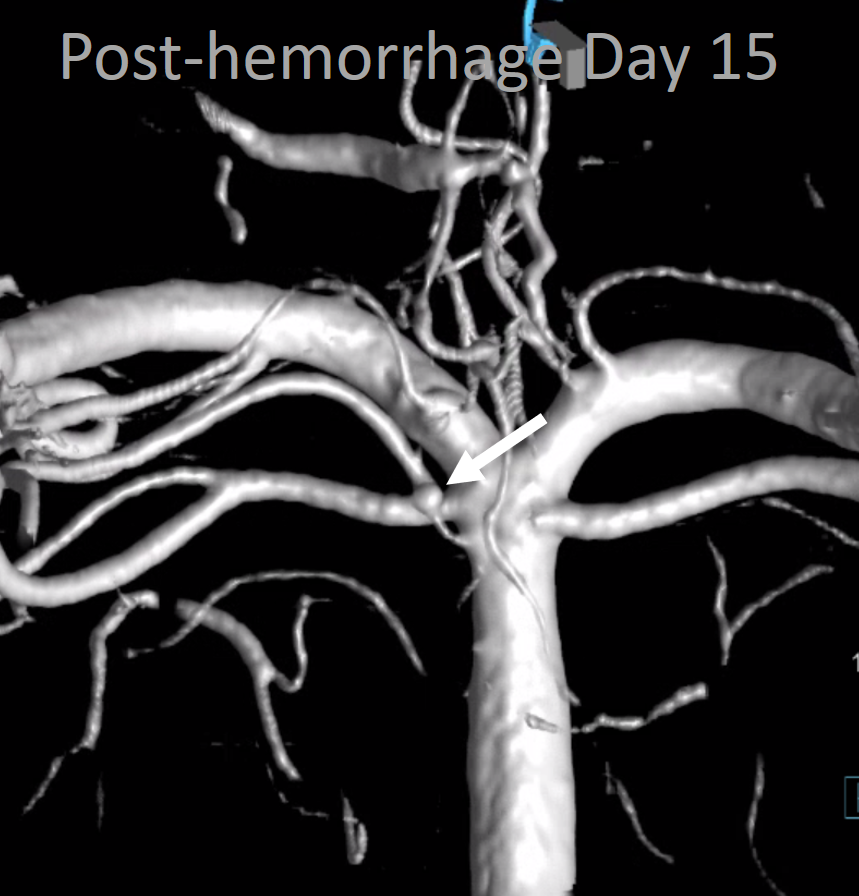
In this patient, a basilar fenestration is associated with an AVM:
A few years later, the AVM is gone, and the fenestration can be better appreciated thru lack of competing inflow from the right vert.
Very rarely, fenestrations can be associated with aneurysm formation — here treated by stent-coiling following rupture. Case courtesy Dr. Moses Aronov. Instragram @neurointervention_Moscow
While the fenestration might be sporadic, one might also postulate that dual channels persisted secondary to AVM-related flow demand during fetal period. This is a tempting, though likely erroneous assumption. Here is another one, courtesy of Dr. Daniel Sahlein:
An ex vivo specimen, courtesy of Peggy Mason, PhD, Department of Neurobiology, University of Chicago, Chicago, IL. Notice also a nice anterior spinal artery (yellow arrow). Also to be seen are petrosal vein, vein of the lateral recess, and nice pontine perforators.
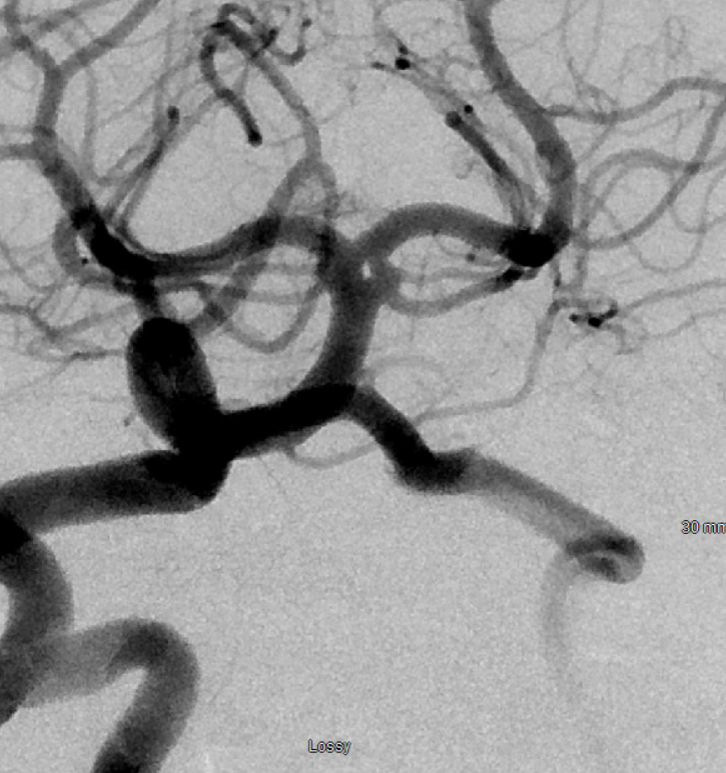
Good angiographic technique is important — the patient below was thought to have an SCA aneurysm
Poor right vert injection, with admixture of unopacified contrast from a co-dominant left vert is not diagnostic — or shouldn’t be
Rotational angio/ VR reconstructions however suggest that a basilar terminus fenestration might be present instead
This requires confirmation — there can be artifacts due to unopacified flow, etc. Optimal projection, strong injection to transiently overcome left vert inflow, and good magnification with no movement confirm presence of this unusual fenestration. See full case here
Finally, a double fenestration courtesy Drs. Shepherd and Raz
Mid-Basilar Hypoplasia
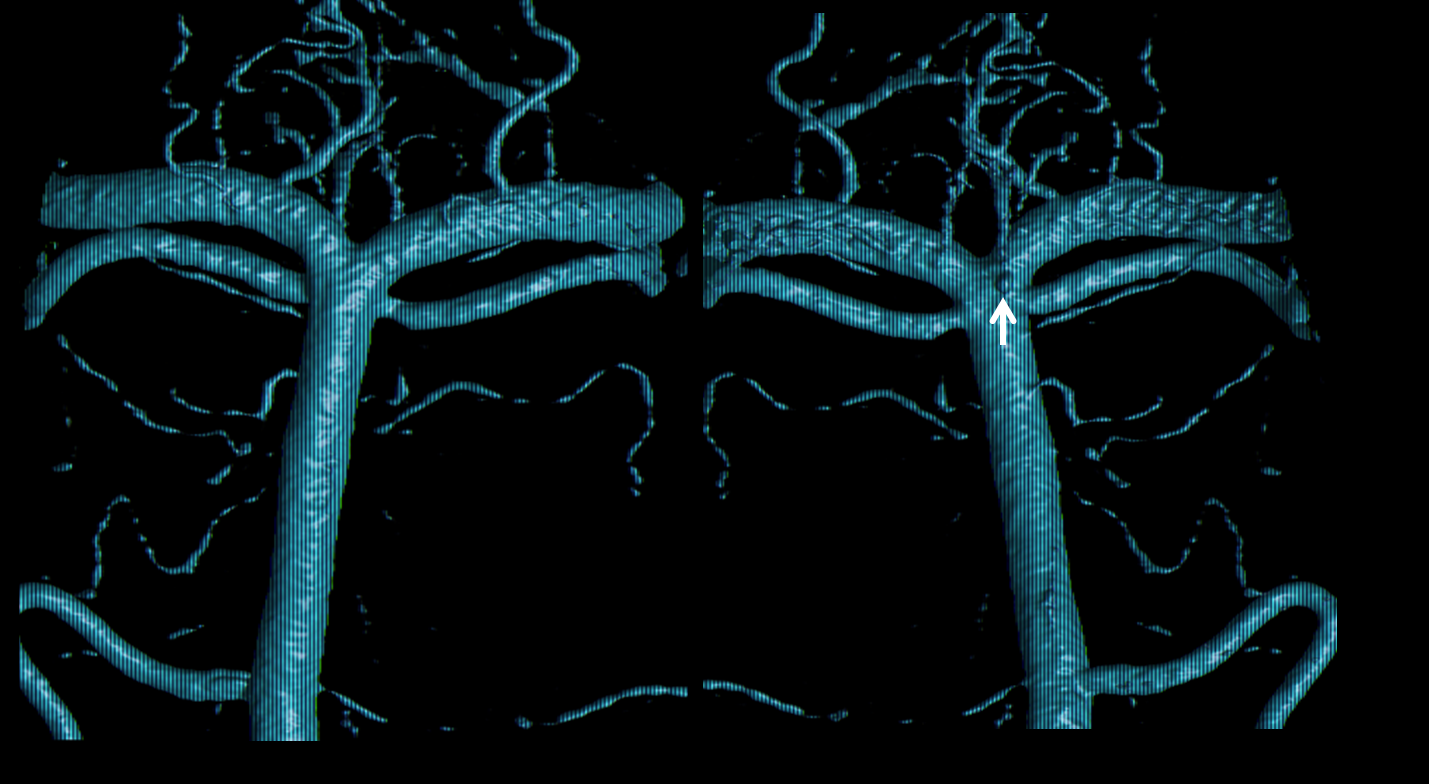
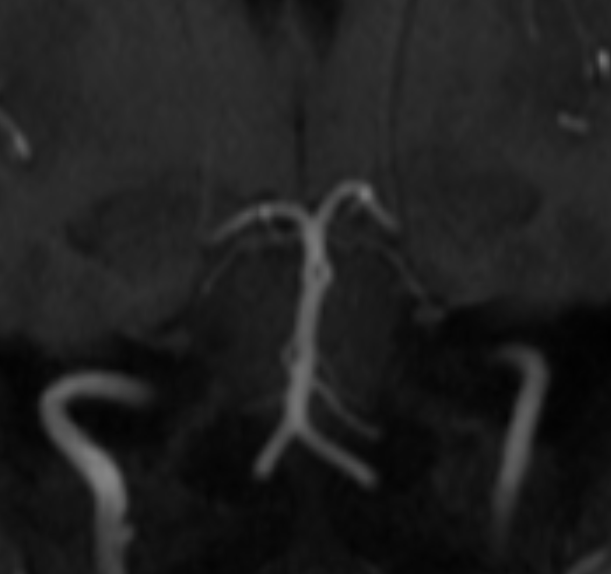
An important fairly rare consequence of embryology is mid-basilar hypoplasia. This is different from bilateral fetal PCOMs. In basilar hypoplasia, the SCA territory is supplied via the PCOMs, and a short segment of mid-basilar artery has an extremely small diameter. Differentiation from pathology can be tricky, since atherosclerosis affects the mid-basilar as well. Below is an example of developmental mid-basilar hypoplasia in a young patient with no athero anywhere
Pseudostenosis of the basilar artery — in some degree possibly related to embryologic boundary in the mid-basilar, as above.
This patient underwent an angiogram for evaluation of a midbasilar stenosis as seen on the MRA. No stenosis was found. This is occasionally seen with time-of-flight MRA imaging — a flow-based technique. If flow is diminished in a segment of a vessel, an erroneous appearance of stenosis can result. It is particularly common when decent-size posterior communicating arteries are present — thus generously supplementing posterior cerebral and even superior cerebellar arteries, and secondarily diminishing the amount of forward flow in the basilar distal to the AICAs. Even though as a radiologist you have to call them like you see them, reasonable scepticism may be useful here, and a CTA rather than an angiogram can be of value also. Notice also an incidental duplicated left SCA (orange)
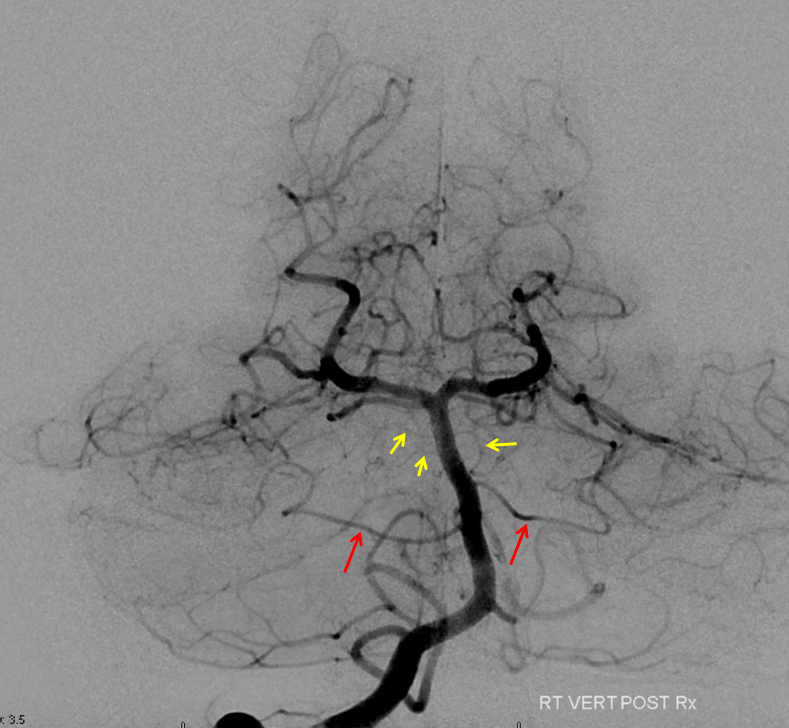
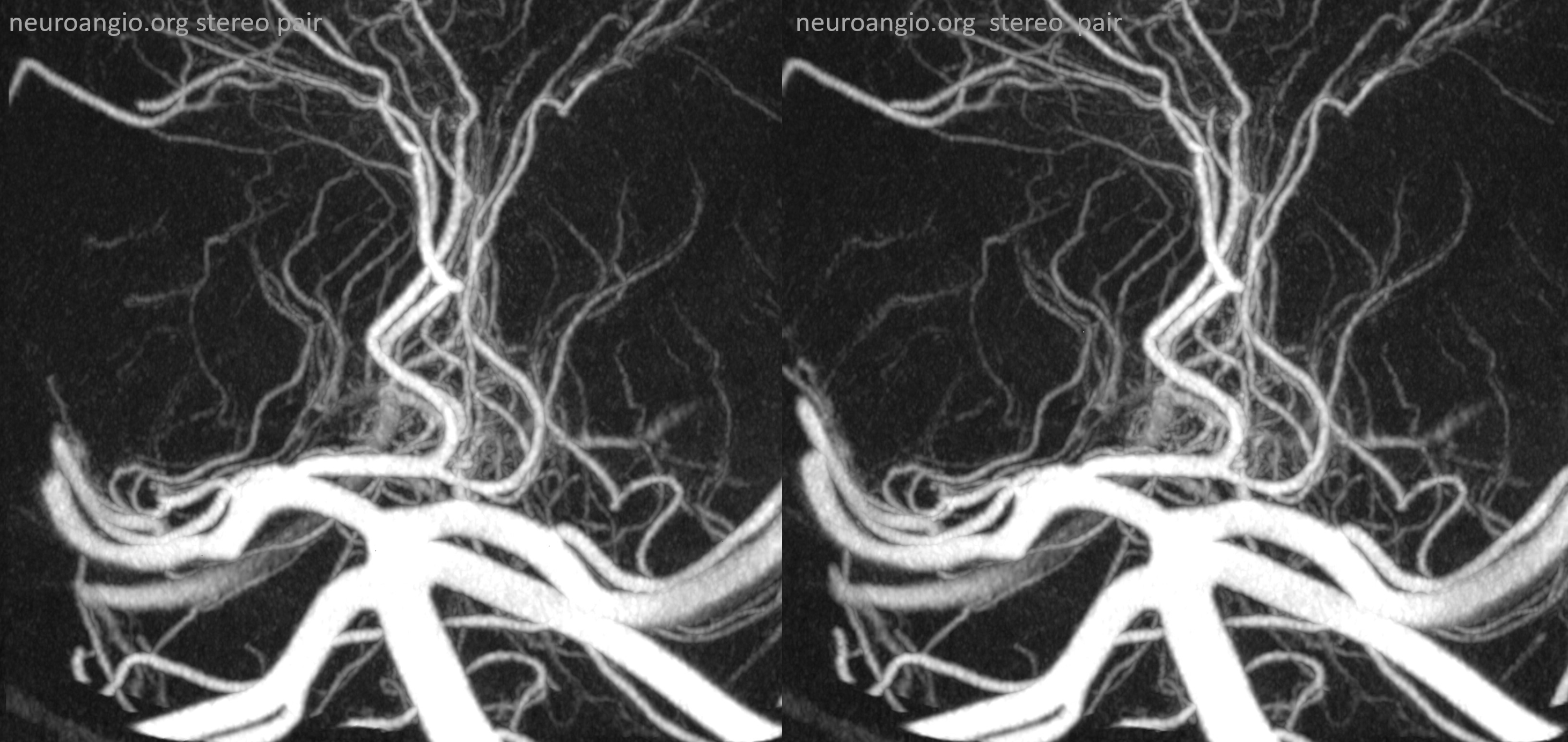
Pontine Perforators:Fronal projection view of the basilar artery, with several transverse pontine “ziggii” perforators (yellow and brown) between the SCA and the duplicated AICAs (red arrows). A loose correlation exists between the number and size of these perforators and distance between the SCA and the AICAs — the longer the distance, the more and larger the perforators tend to be.
Submental view of the basilar artery (below), showing different positions of left left and right AICA (red arrow) origins — again supporting the notion that AICAs develop from a number of possible choices — representing basilar artery perforators. Notice also the perforators themselves (yellow arrows), of which there are two visible ones on the right (likely because of larger distance between SCA and AICA) and one on the left (shorter SCA/AICA distance). Both AICAs are about equal in size, vis a vis AICA/PICA balance. The “Post Rx” refers to stenting of right superior cervical vert dissection/pseudoaneurysm, below the field of view.
Diagram of the same disposition (right image), depicting origin of the AICAs from a somewhat lower right and higher left perforators, as compared with the classical disposition (left image)
High-riding basilar — the position of the basilar tip (red) in relation to the posterior clinoid (white) and less importantly to the petrous apex (blue) is critical in pre-surgical assessment for those who still clip basilar tip aneurysms. An aneurysm neck below the posterior clinoid tip is difficult or impossible to reach and therefor to adequately clip even with a subtemporal appoach. This is a high-riding basilar. Also notice downward sweep of the PCAs relative to basilar tip — another indication of high basilar positon. The opposite istrue of the low basilar
Upper Basilar Perforators
There is a wealth of very clinically significant perforators arising from basilar bifurcation and proximal P1 segment, usually supplying the mesial thalamic structures. The classic “Percheron” is a common origin of such perforator supplying bilateral thalami. Clinically important as injury / occlusion leads to profound level of consciousness alterations.
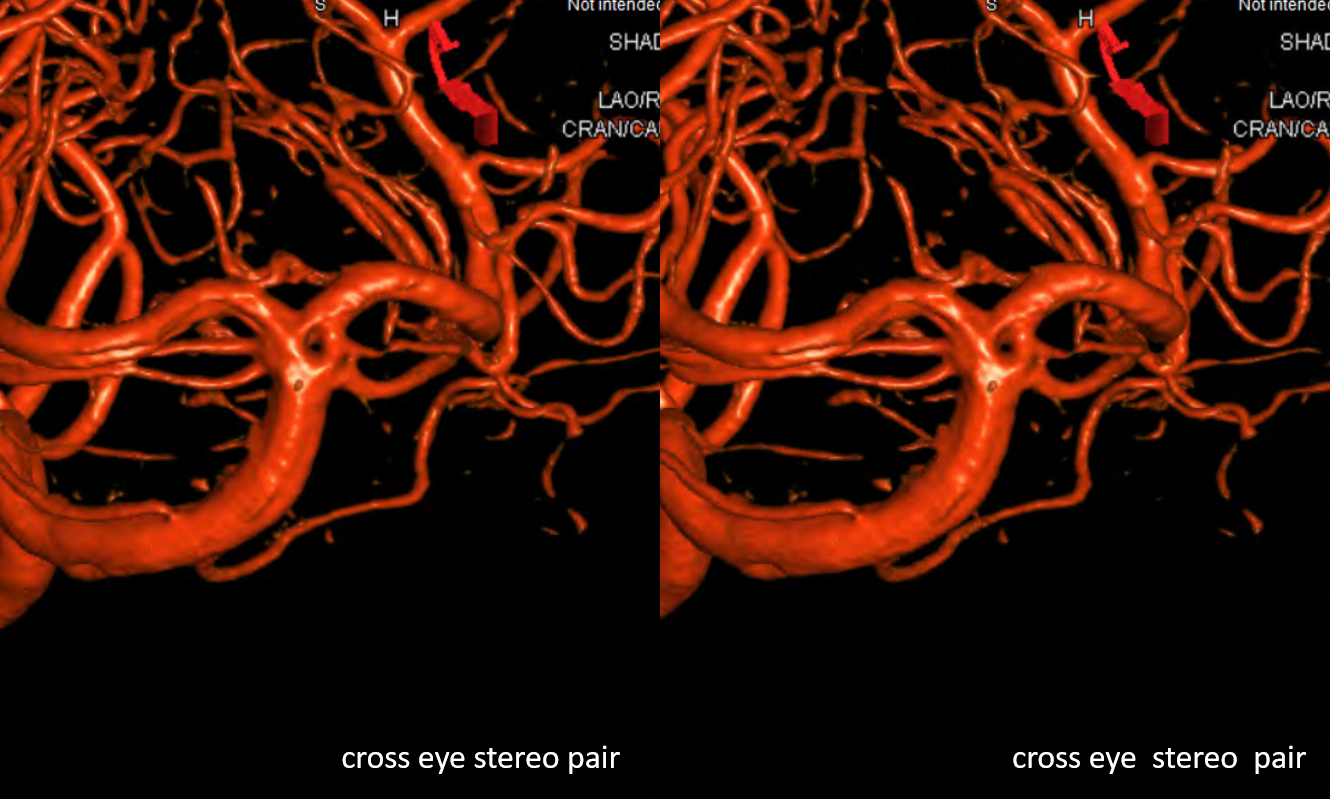
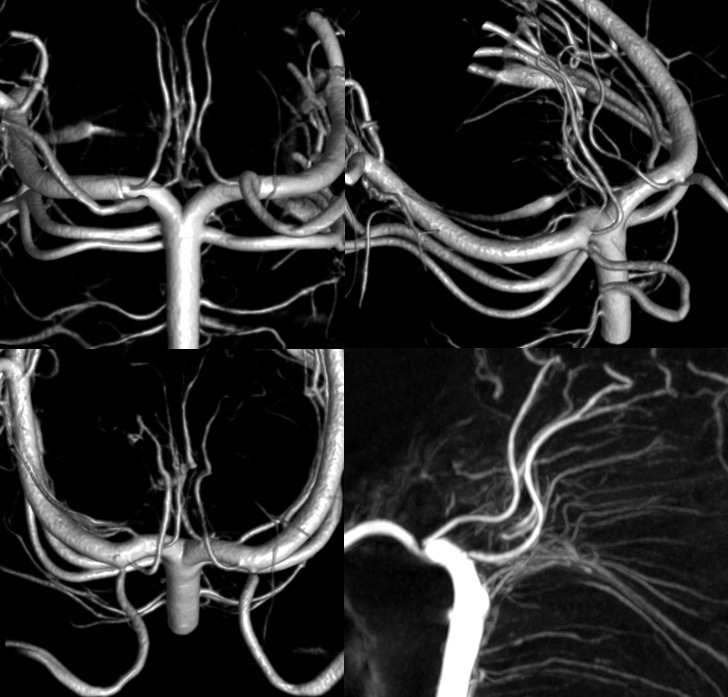
As in anything — there is a spectrum. One might have a common lenticulostriate group origin or individual ones. Below is a beautiful set of DYNA VR images by Eytan Raz. Note how the more lateral pair of perforators is also more anterior — as is in balance with anterior thalamic perforators typically arising from proximal PCOMs. Also present is a “duplicated SCA” and bilateral “duplicated” AICAs. All part of the same big spectrum idea
Percheron Stereo
Another Percheron Stereo
Another Percheron DYNA
In addition to percheron (dashed arrows), one sees bilateral paired long collicular arteries (arrows) and PCOMs
The medial / posterior thalamic perforators tend to originate from the basilar even in bilateral fetal PCOM cases. These are rare — at least having no P1 at all is much more uncommon than having no PCOM. I does present a bit of a conundrum from embryologic perspective, since the “fetal” PCOM is supposed to be the early way — having more nonexistent PCOMs than P1s is a bit of a problem with this theory. But that is besides the point. The Clinical issue is that these perforators will come off basilar even when P1s are absent — see below
Basilar Occlusion Syndromes
These span the spectrum between asymptomatic and fatal. Focal mid-basilar atheromatous occlusions are extremely well-tolerated when PCOMs are present to supply what’s above. When absent, result is usually death or major disability.
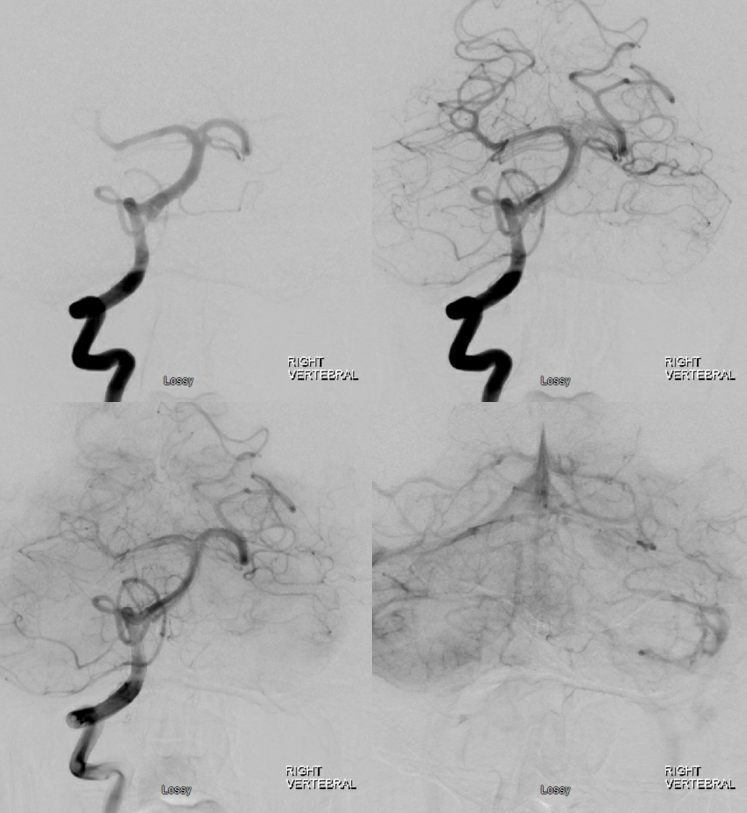
Long segments of basilar occlusion, without PCOMs, are equally deadly. With PCOMs, the result is usually pontine perforator infracts. Unilateral single perforator infarcts have good to excellent prognosis. Bilateral infarcts of multiple perforators, while sparing top of basilar, produce the dreaded locked-in syndrome. See below. Initial presentation was right-sided weakness, due to larger burden of occlusions on the right side, progressing quickly to full syndrome.
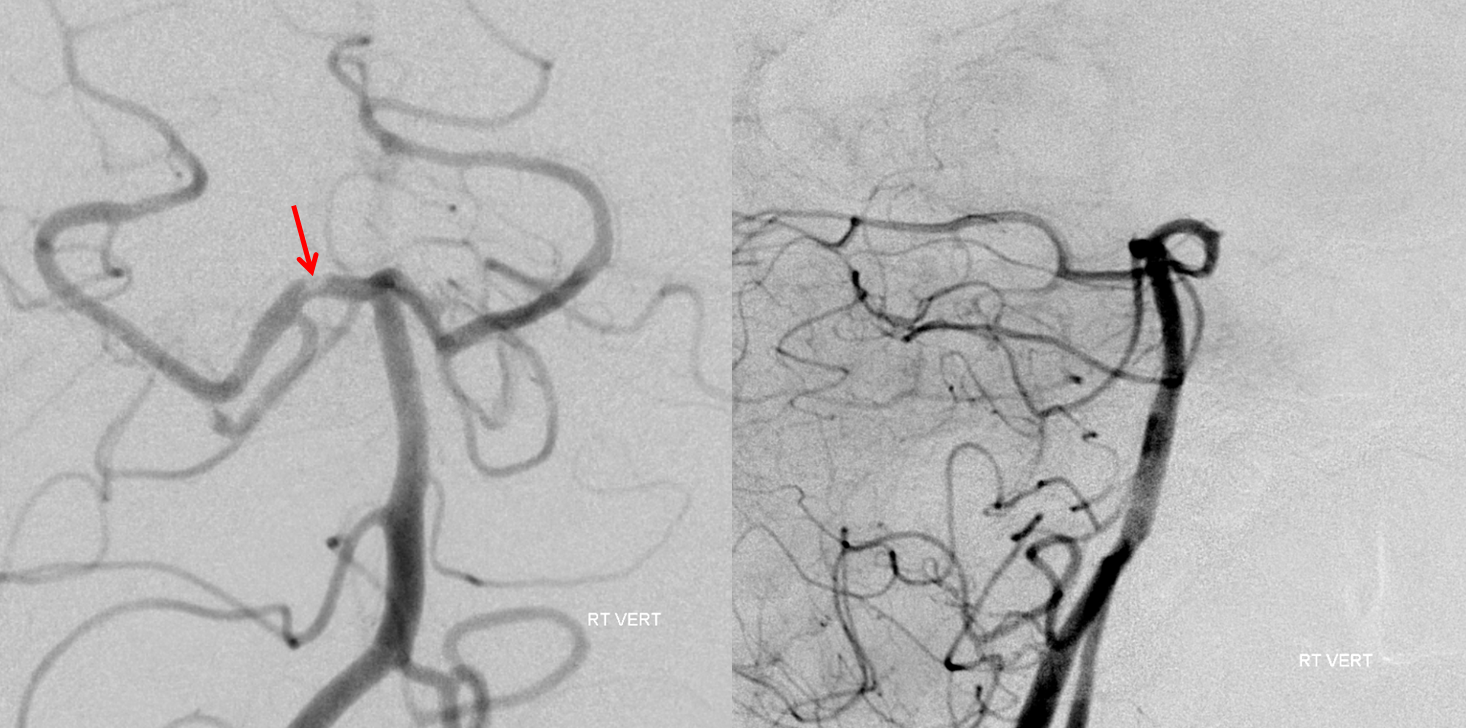
Focal top of basilar occlusions, when blocking thalamic perforators, result in profound decline in consciousness, often markedly out of proportion to size of embolus. Here is a subocclusive embolus in a comatose patient. Notice lack of thalamic perforators on both images
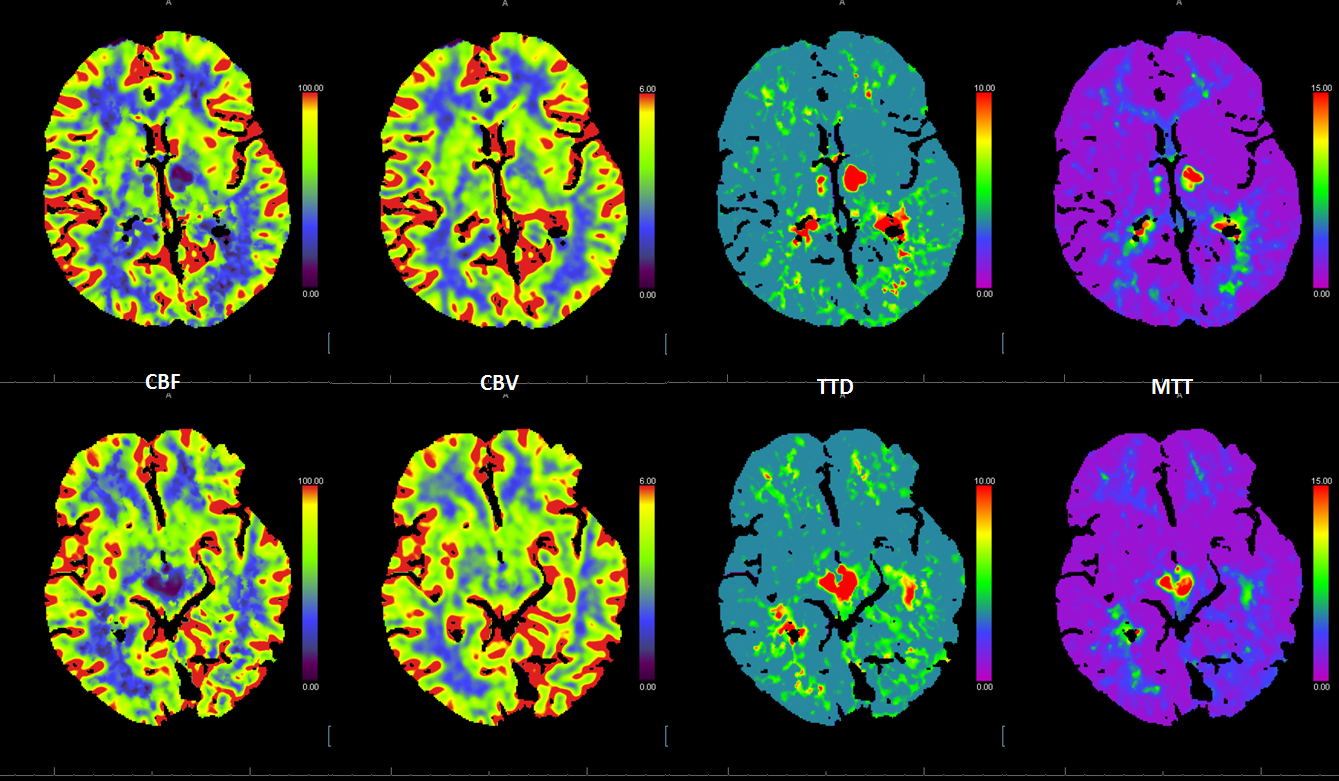
The perfusion images below are actually the most informative — everything is there. it is one of the most amazing perfusion images we ever came across.
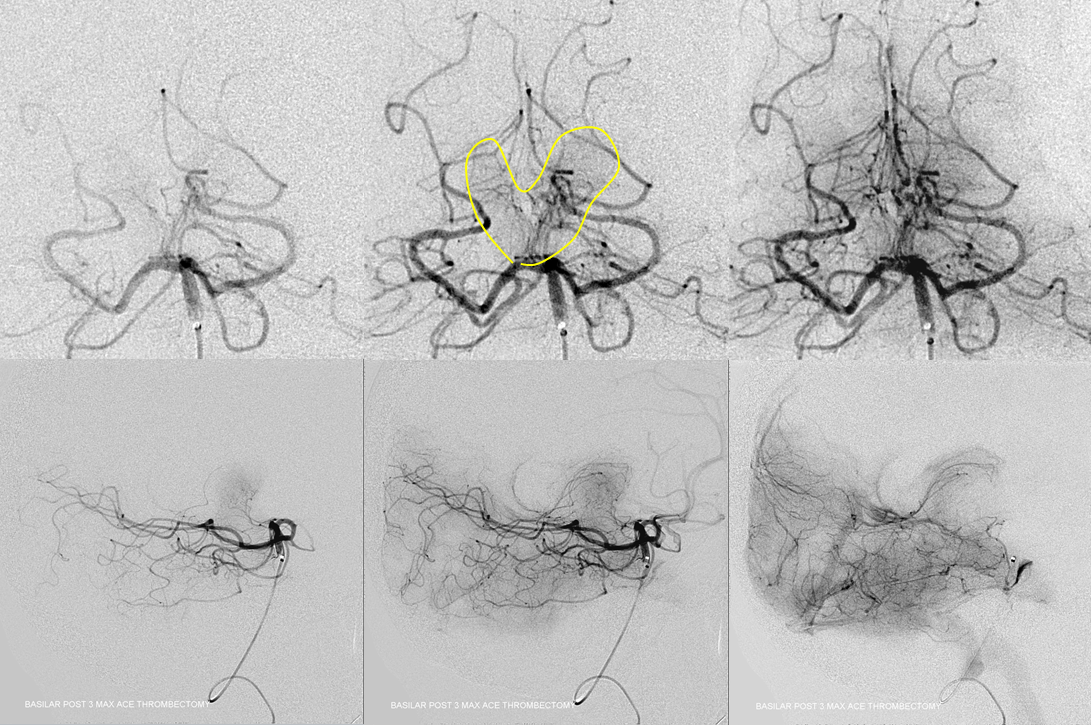
Post thrombectomy — all thalamoperforators are arising from the site of prior occlusion…
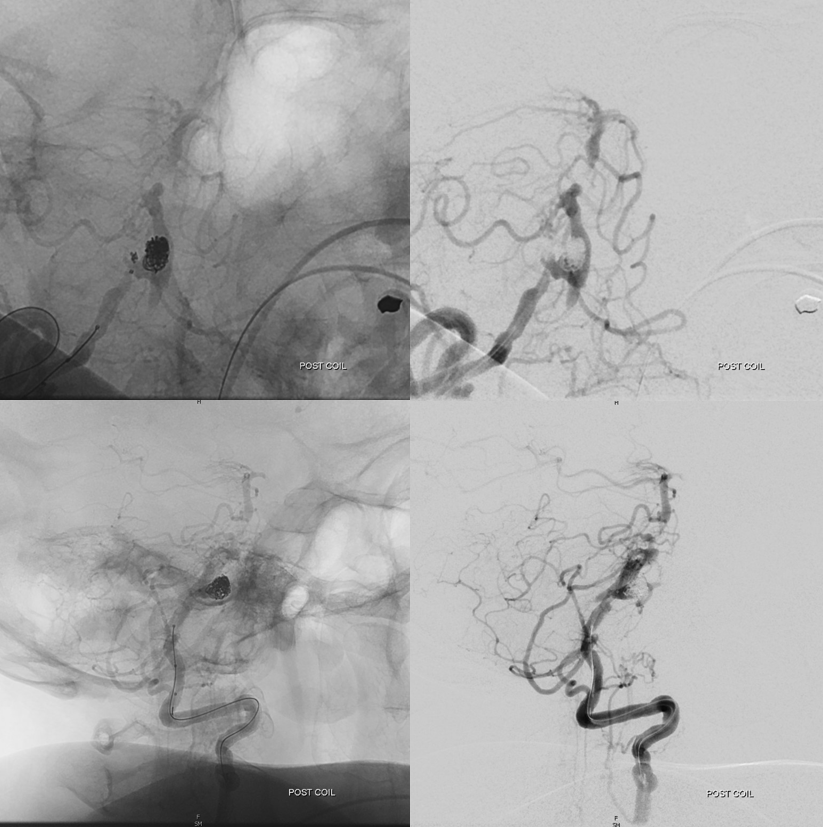
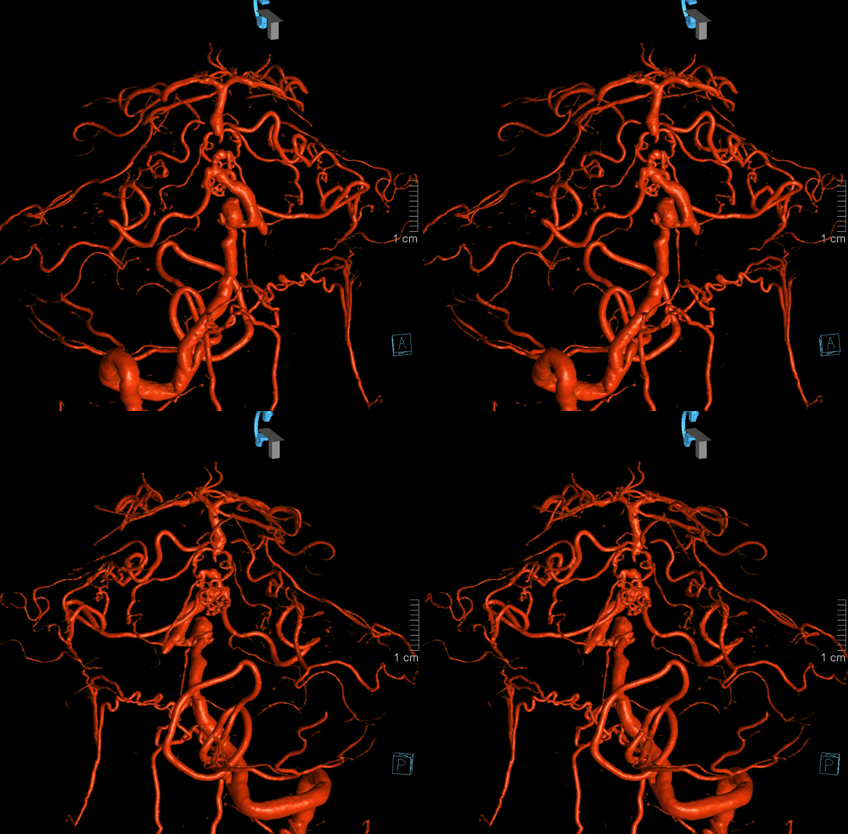
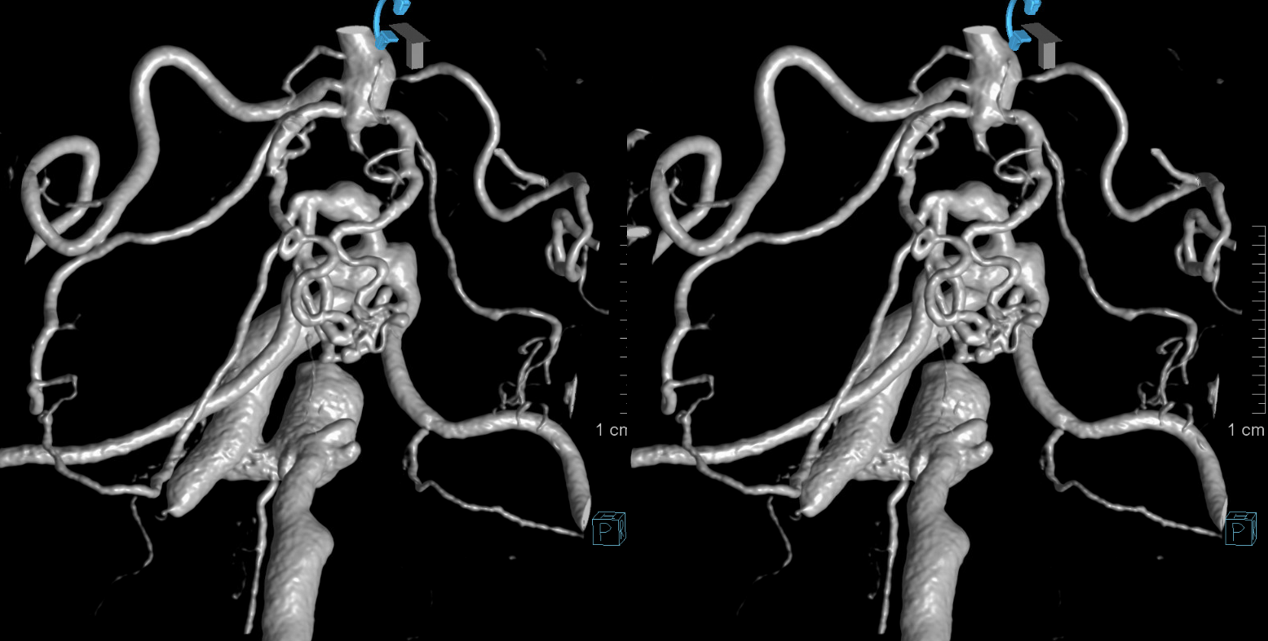
As in many slowly progressive steno-occlusive lesions, extensive collaterals can develop. Focal mid-basilar occlusion can be extremely well tolerated — asymptomatic, minimally so, and pure motor syndrome making excellent recovery — typically when good PCOM-P1 connections supply the upper circulation. When PCOMs are absent, occlusions can be devastating. Or not — if regional collaterals do their job. Below is an example of multiple AICA-SCA and regional mid-basilar perforator collaterals in a patient presenting with SAH from a ruptured aneurysm. Prior to rupture patient was asymptomatic
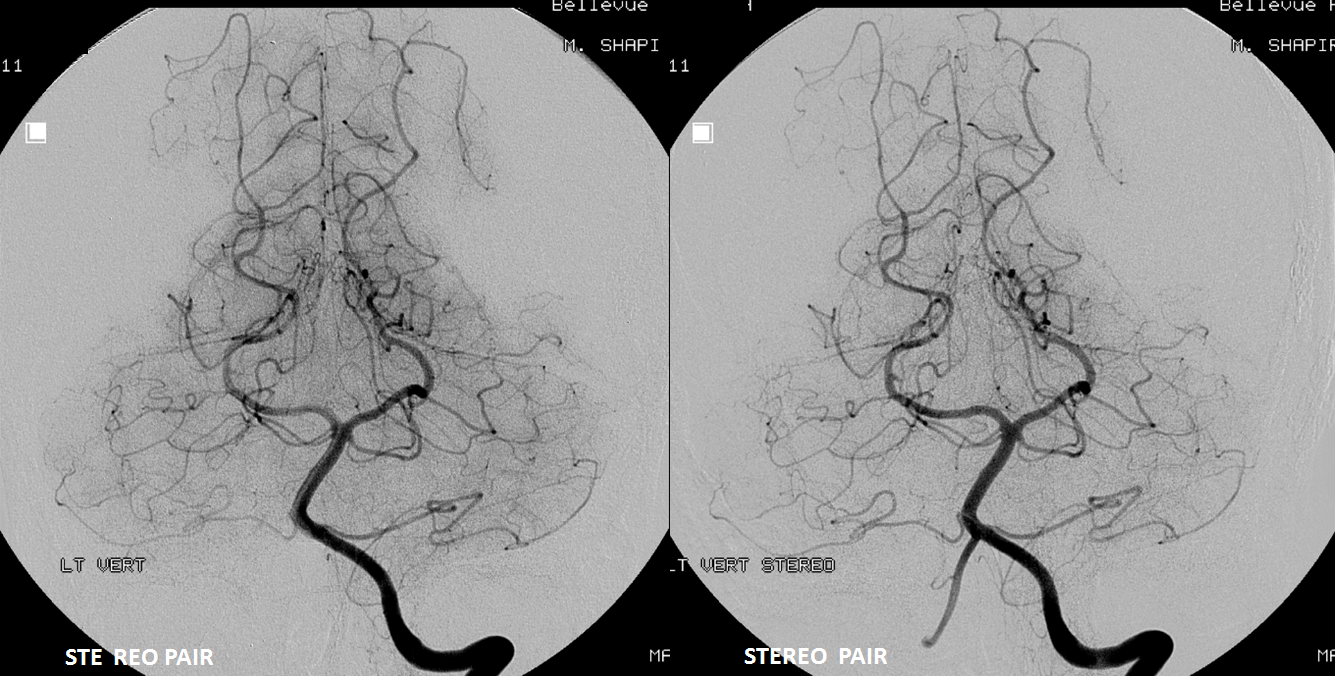
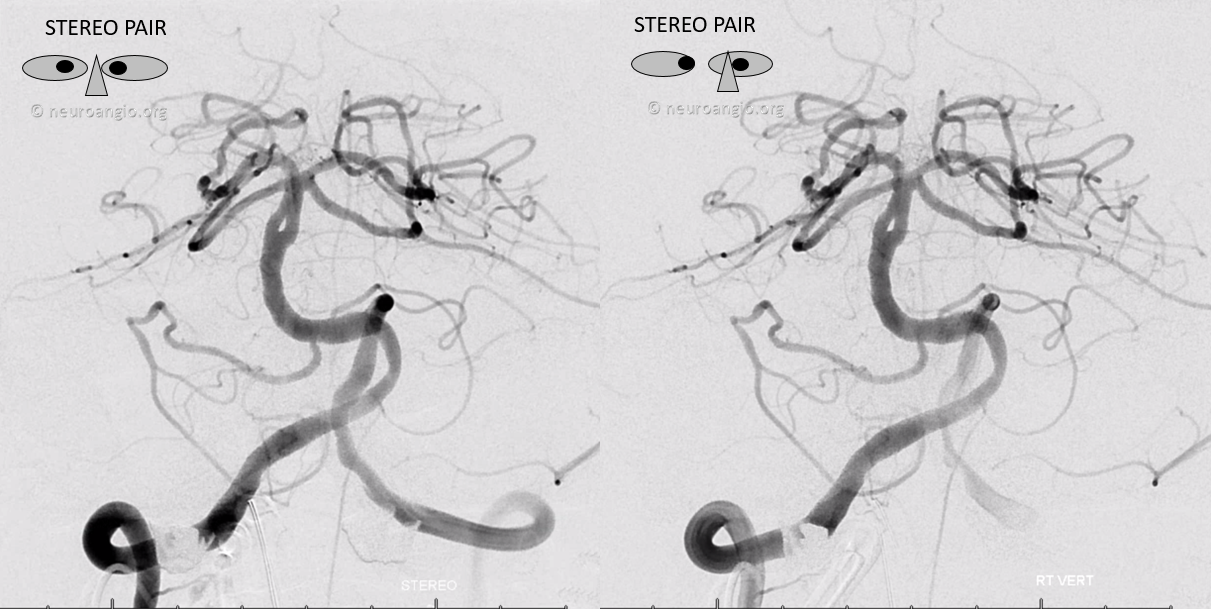
Stereo pairs below — every collateral route is present. There are hemispheric AICA-SCA ones laterally. Then the Primitive Basivertebral Anastomses of Padget more medially. And, finally, proximal parasagittal anastomoses between regional mid-basilar perforators. The paired nature of mid-basilar perforators is perfectly accentuated here by the paired enlarged anastomoses. Can you see them all?