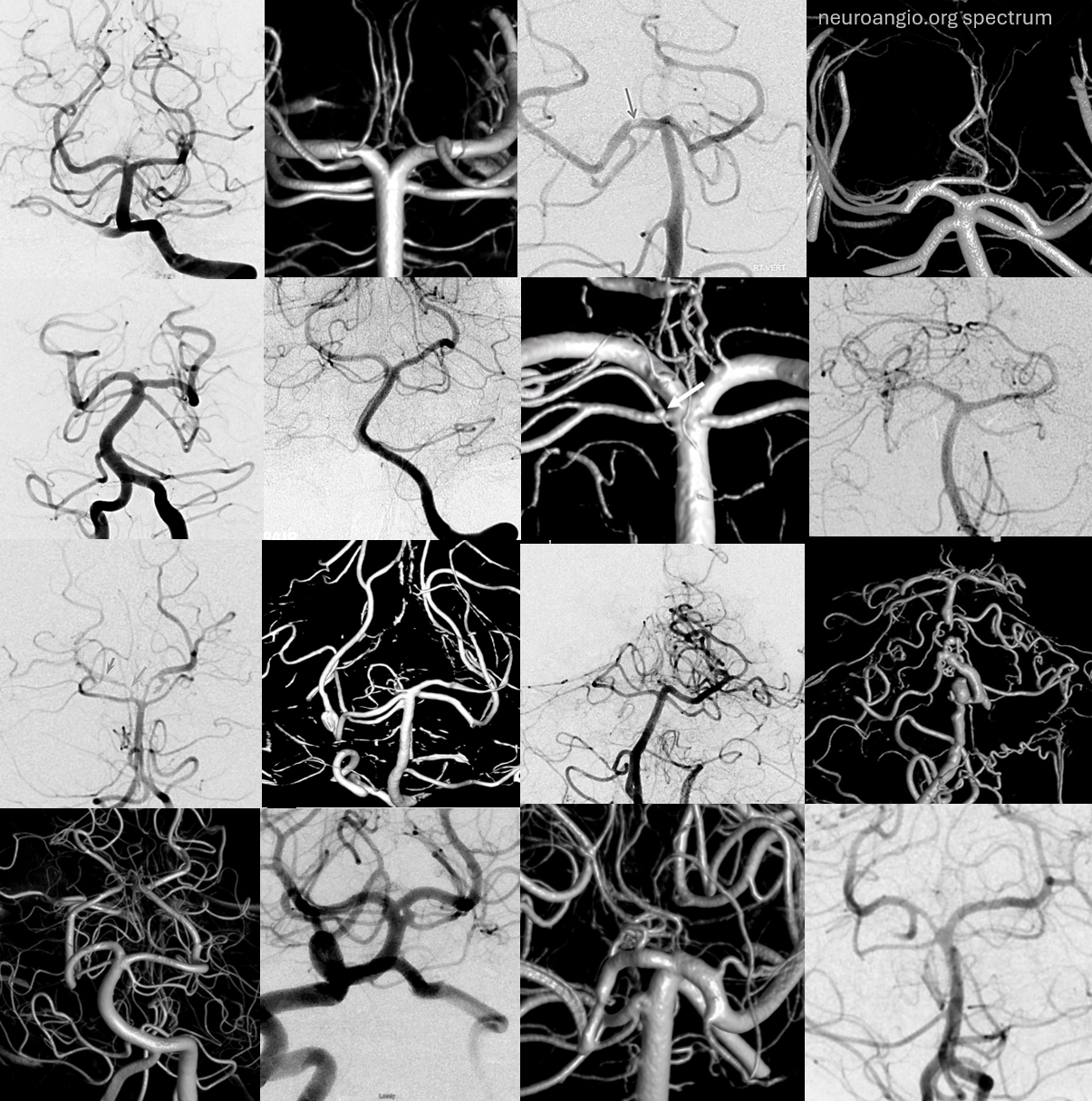
The collage below shows the wide spectrum in SCA branching patterns and the futility of assigning these endless variations anything but a very qualitative description
The superior cerebellar artery earns a distinction in being the most constantly identified vessel arising from the basilar, probably because its territory does not lend itself to the same degree of plasticity (with respect to arterial supply) as do those of AICA and PICA. In contrast to the latter two vessels, whose size and territory are inversely proportional to one another, the territory and, therefore, size of SCA is more constant. It is also phylogenetically older than the PICA, which was “impressed” into cerebellar service at a later time, and thus continues to wrestle with the AICA to define its territory. The said territory of the SCA, classically, consists of the superior pons and pontomedullary junction, superior cerebellar surface, much of cerebellar white matter, dentate nucleus, and superior vermis. While invariably present, the SCA is duplicated in at least 25% of subjects. (See below for discussion of basilar perforators). When so, the upper branch typically supplies the vermis, and the lower one is hemispheric. When not duplicated, the vermian branch will often come off very proximally, so as to be “almost” duplicated. Often, one vermian branch is present only, as it is an unpaired structure. There are, of course, named segments of the SCA, could it be otherwise? They are prepontine, ambien, and quadrigeminal — and useful in terms of relating the artery to its landmarks and, therefore, territories of supply. Usually, quadrigeminal segment is responsible for the inferior colliculi only.
“Classical” view of the SCA, originating from rostral basilar artery just below the basilar bifurcation.
Vermian and Hemispheric Divisions
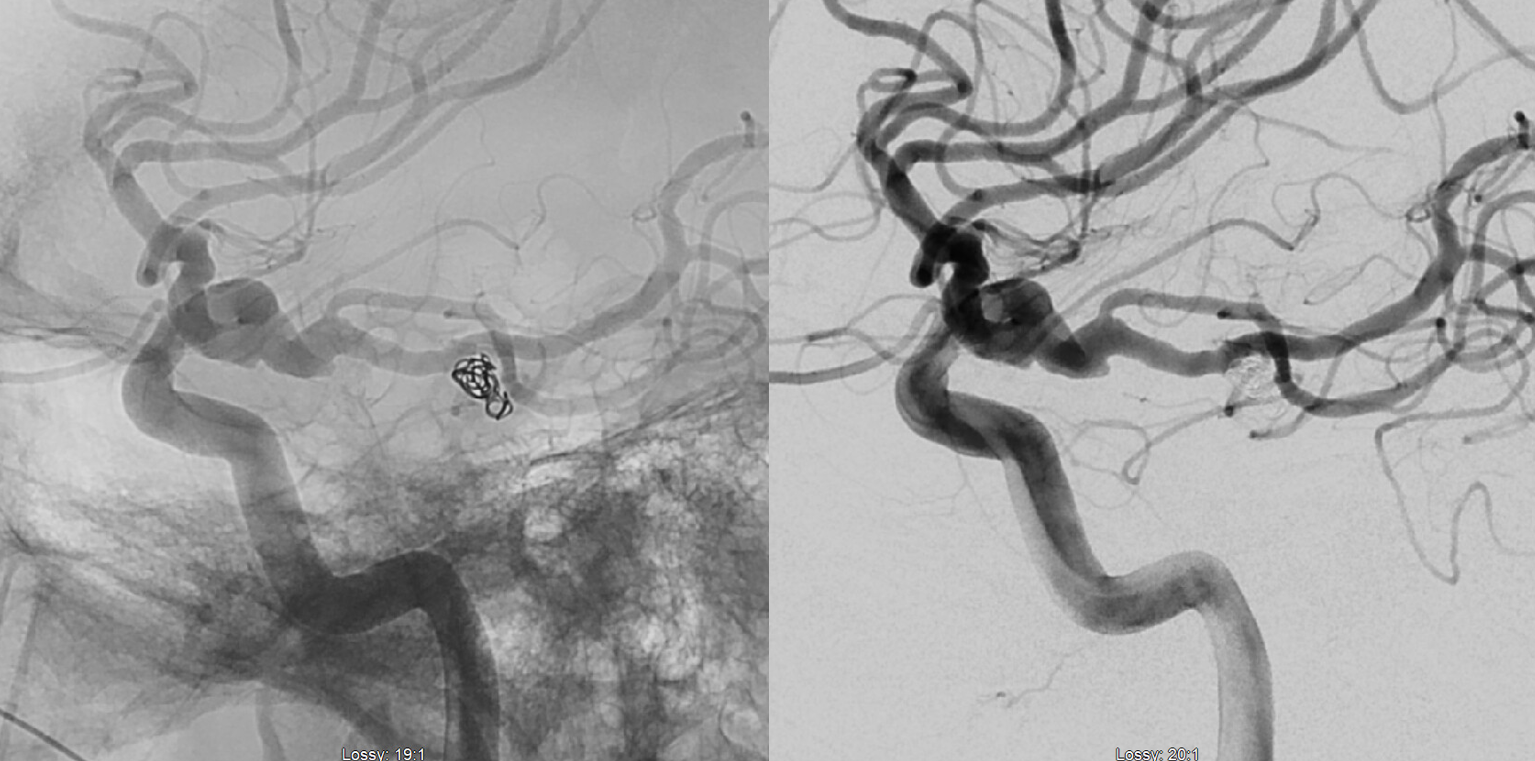
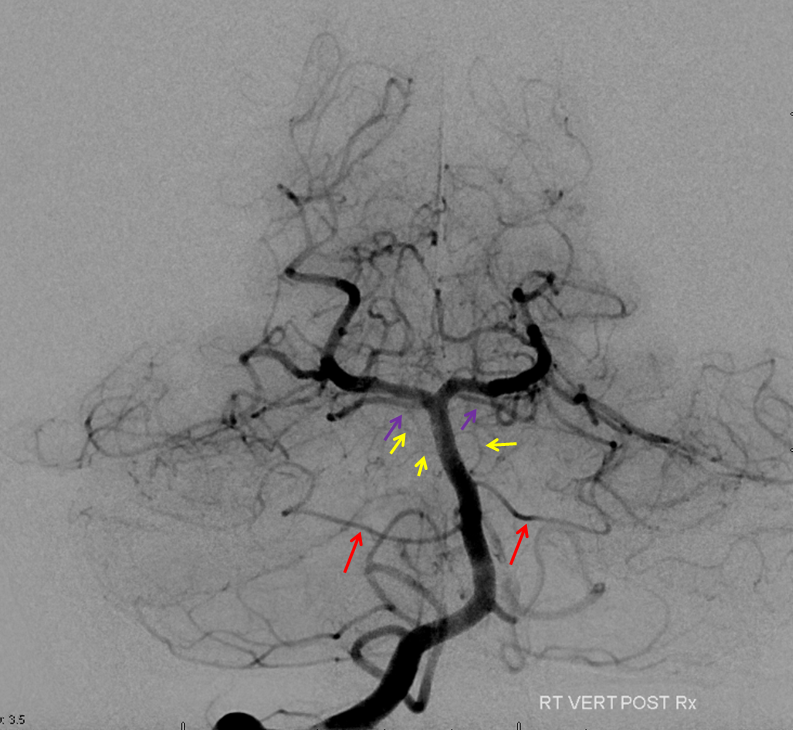
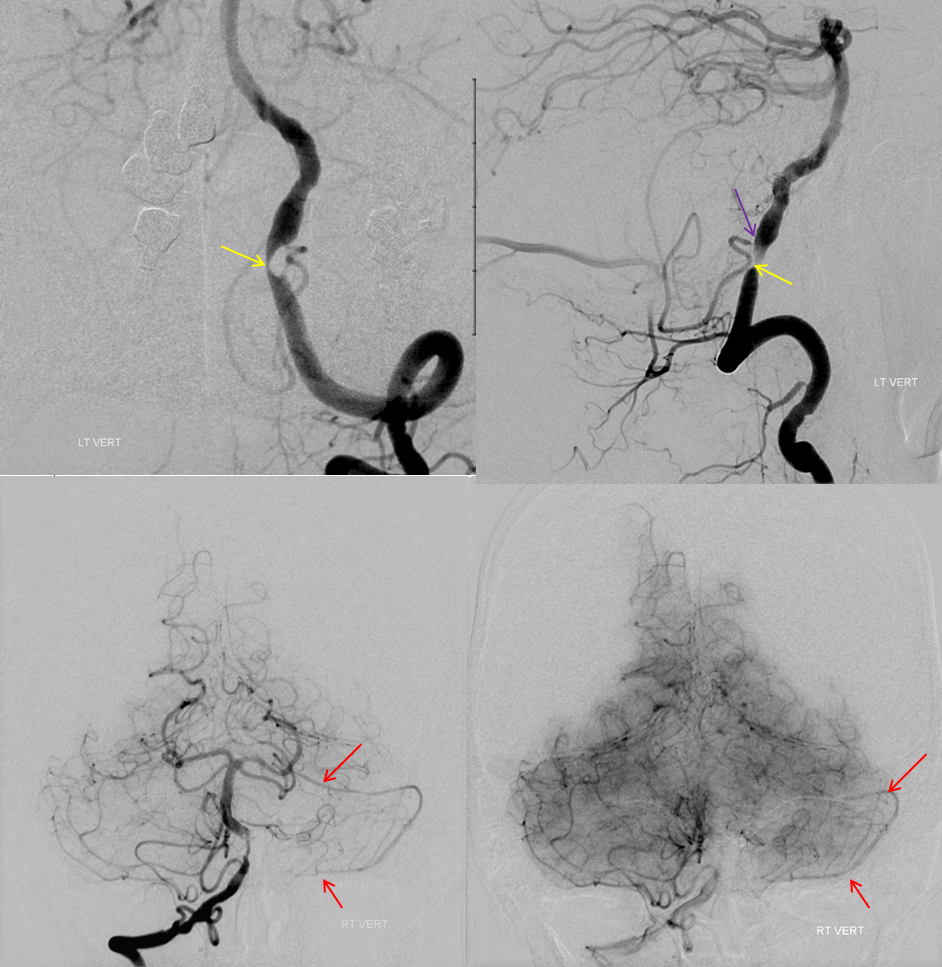
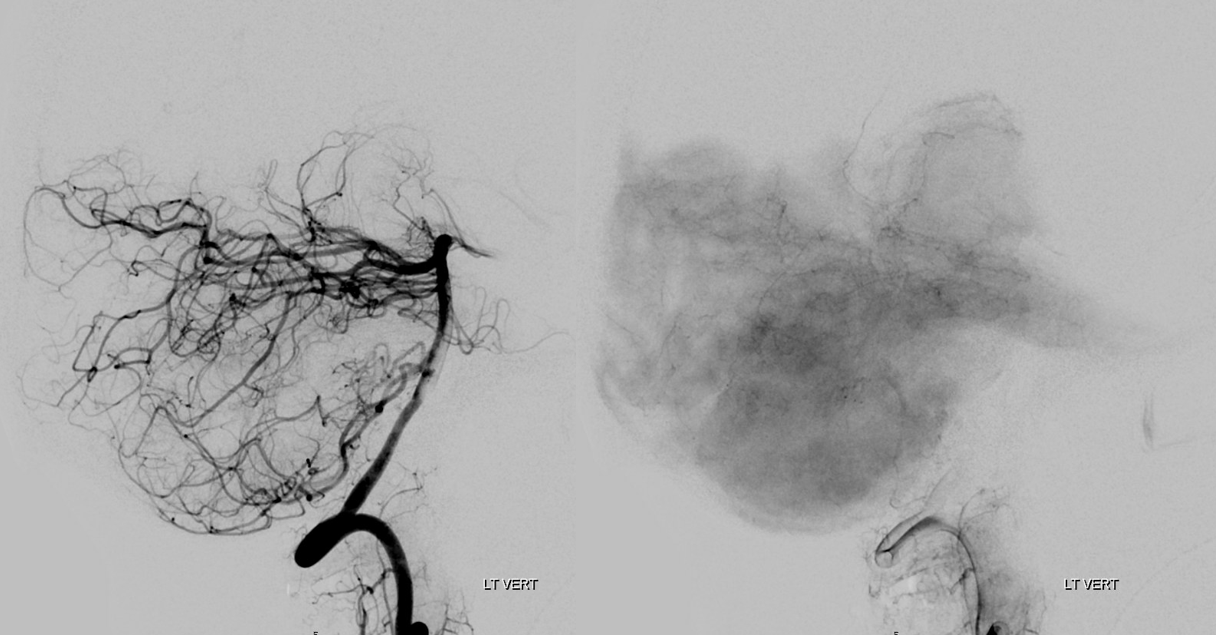
There are basically two divisions. The bifurcation is extremely variable — from very distal to “duplicated” — arising separately from the basilar artery — SPECTRUM again. The vermian branch is almost always the upper one — either more distal when arising from basilar trunk or the upper one when bifurcating from single branch. Below is an example of an SCA fusiform aneurysm. On the right separate origins of vermian (cranial) and hemispheric (caudal) branches. On the left single branch, with upper one being vermian. Best seen in upper right and lower post Rx images. Full case here
Variation in fusion patterns of the basilar artery, as pertaining to SCA
To a large extent, variations in SCA disposition are discussed in the Basilar Artery section, which is a necessary redundance. The following discussion of SCA origin is essentially copied from the Basilar Artery section, which should be consulted for the “bigger picture” of the vertebrobasilar system, together with the AICA and PICA sections.
The basilar artery is formed via coalescence of multiple channels belonging to the longitudinal neural system, following development of the anterior circulation (see Neurovascular Evolution and Vascular Neuroembryology sections) This process sets the stage for multiple basilar artery variations, which can be considered from the standpoint of 1) extent of fusion 2) position of fusion with respect to the brainstem, and 3) completeness of fusion (i.e. fenestrations).
It is conceptually helpful to think of the basilar artery as being “zipped” in the middle, with vertebral and PCA segments being “unzipped”. The length of the “zipped” segment, and the integrity of the zipper determine the final configuration of the artery. As such, the artery may be
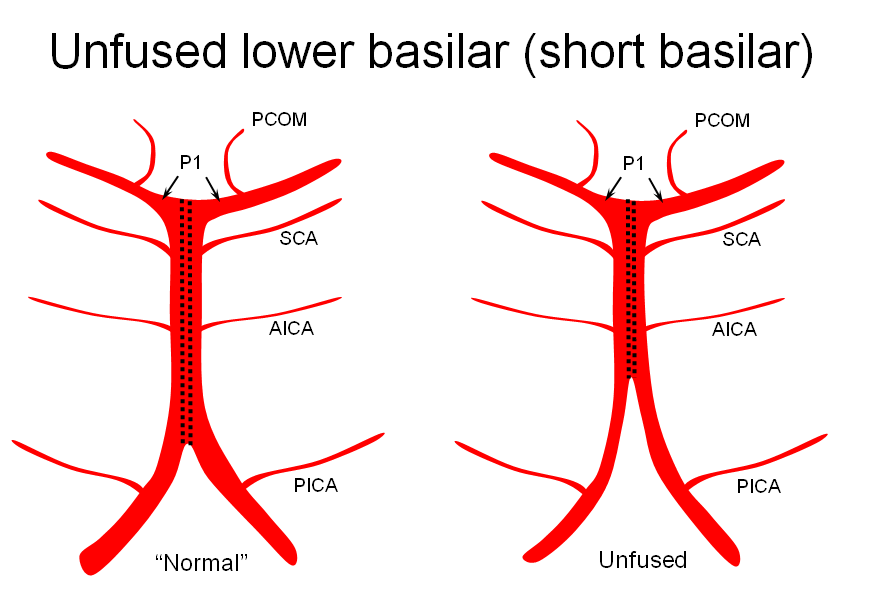
1) “short” — corresponding to relative lack of fusion of the caudal or rostral segments
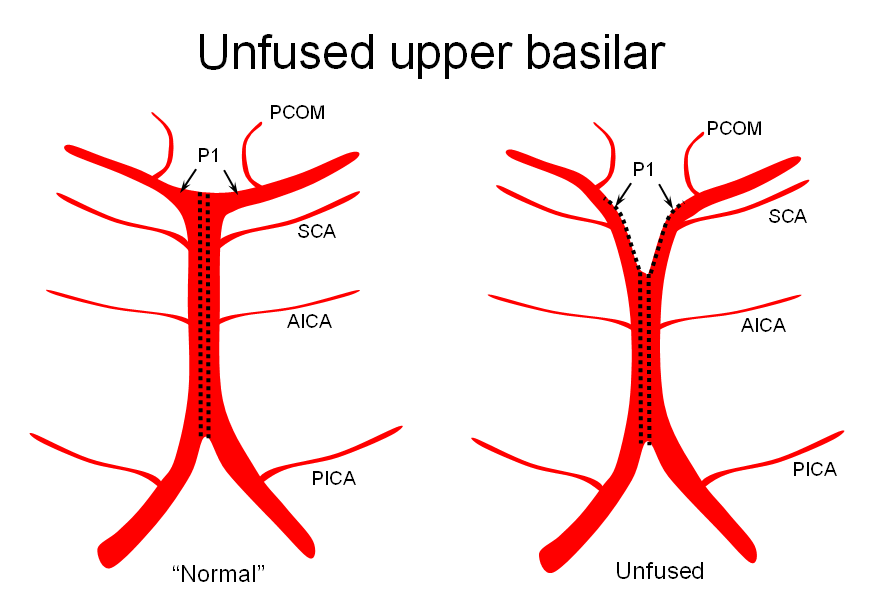
2) Unfused at the top — corresponding to “unzipping” at the basilar tip — less common but much more confusing
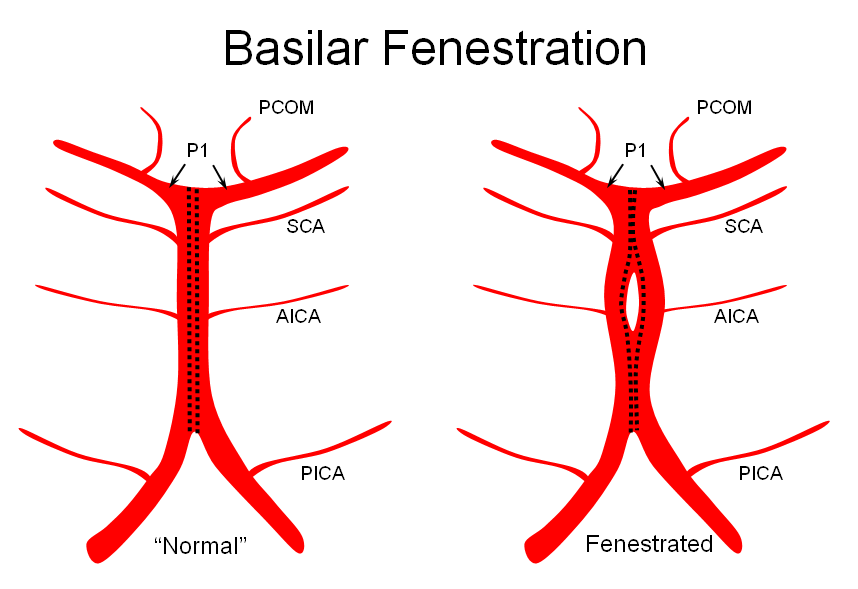
3) Fenestrated — broken zipper in the middle
The first one, short basilar, is in fact a continuum in terms of fusion extent. There is no “number” to guide what short or long is, but some basilars are clearly quite foreshortened. (The tortuous basilar of the old hypertensive is a different matter from developmental variation)
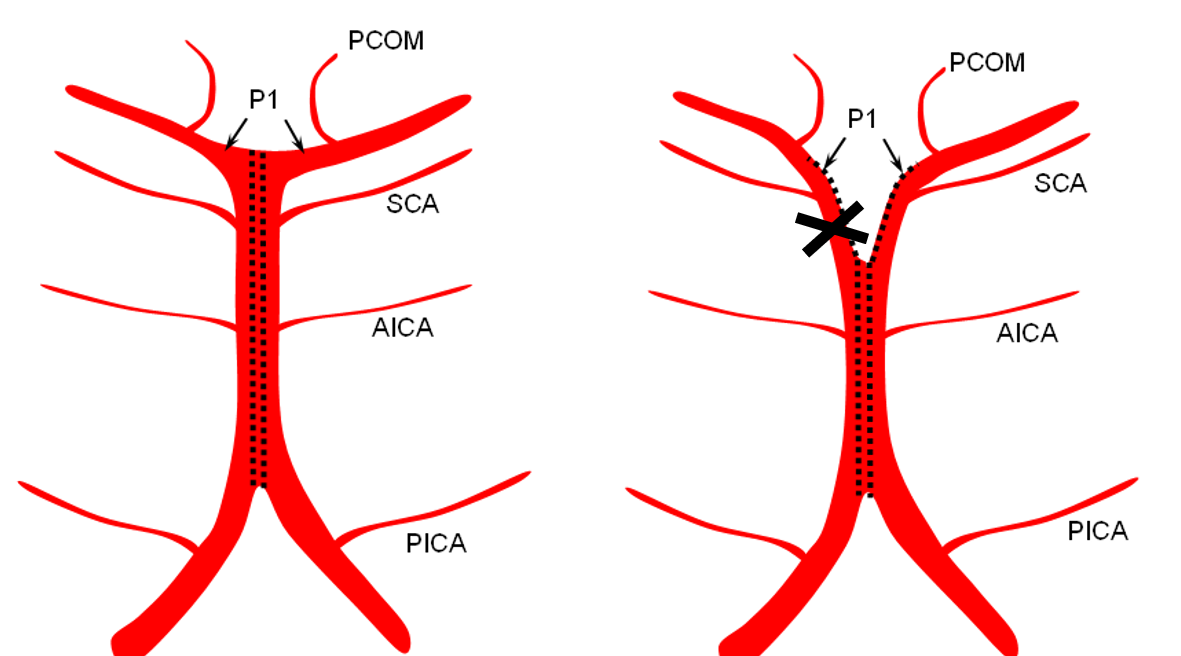
The second variation, involving lack of basilar tip fusion, can generate a lot of confusion. Effectively, the top of the basilar is split in two, so that one or both superior cerebellar arteries originate from the P1 segment. This variant is not, therefore, a primary superior cerebellar artery aberration, but instead a deficiency in basilar fusion.
The third, basilar fenestration, is quite common, and usually of little clinical significance, except when it is so short as to mimic a dissection.
Illustrations
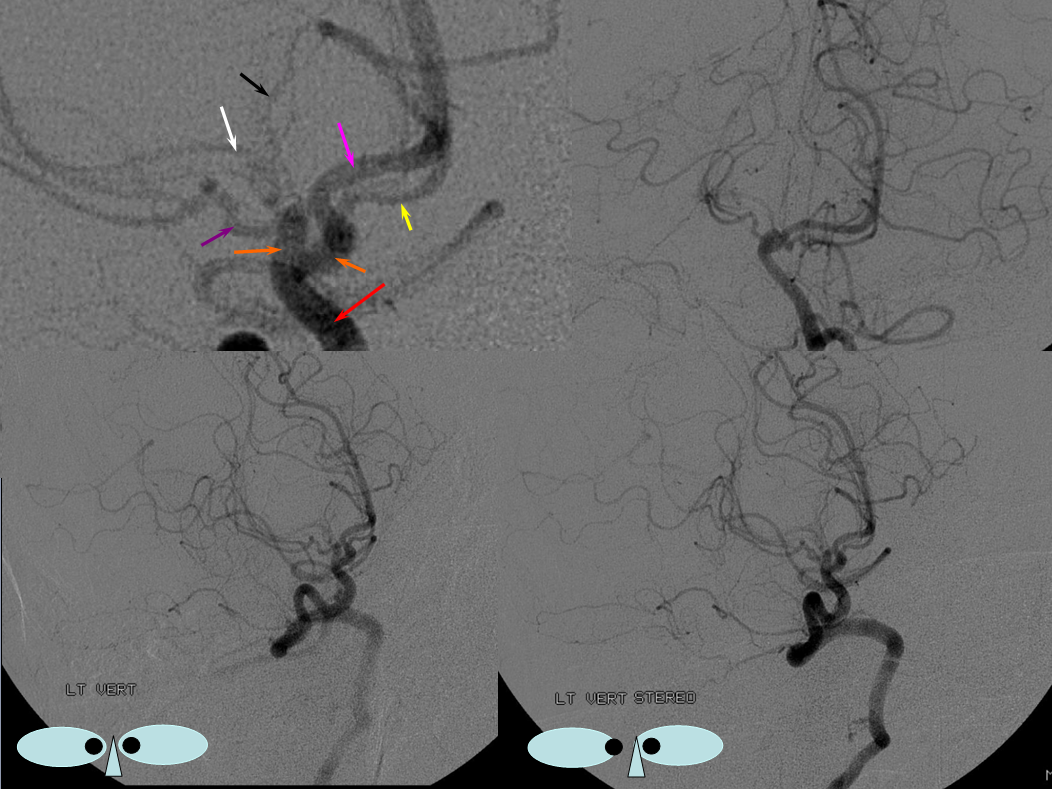
Unfused basilar simulating aneurysm — reported as basilar tip aneurysm on CTA, angio demonstrating an unfused basilar tip with tortuous P1 vessels simulating aneurysm, further complicated by presence of a fetal PCOM on the right.
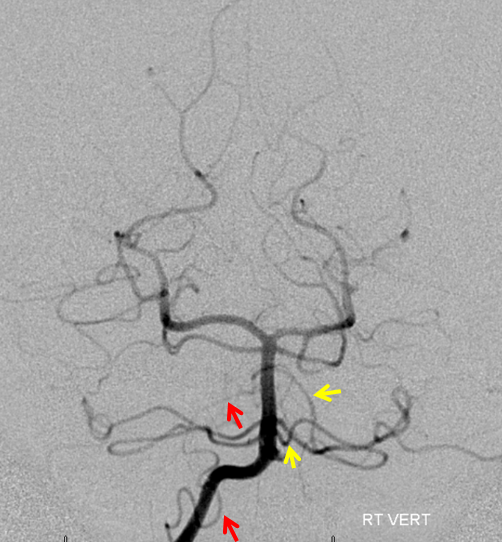
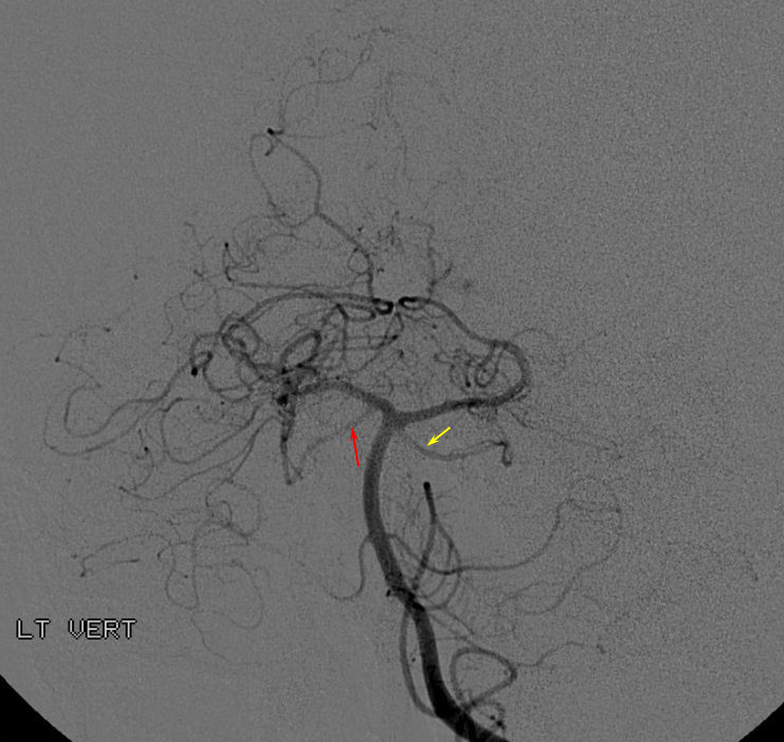
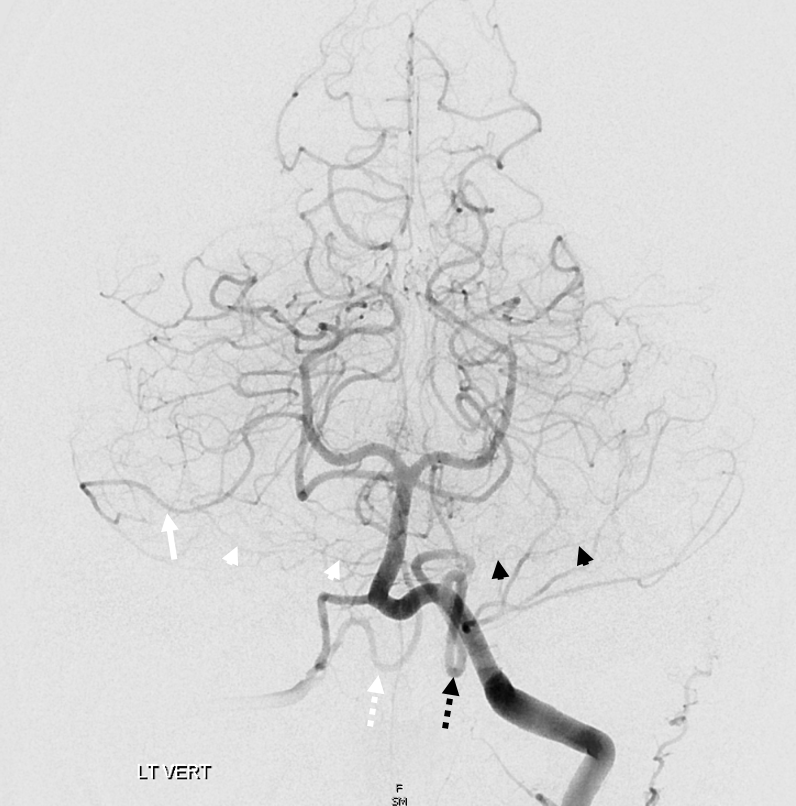
Origin of superior cerebellar artery at “top” of the basilar on the left (yellow), and from P1 segment on the right (red arrow)– seen often, can be conceptualized as a “short” basilar which did not undergo enough coalescence at the top to incorporate the superior cerebellar artery.
Notice P1 origin of the right superior cerebellar artery (red) with contralateral classical disposition in yellow
SCA fenestration — why not?
Below is a very rare situation that Eytan came across, of apparent SCA origin from the PCA, without connection to the basilar artery (marked by X). This might be developmental or acquired — if there is focal athero at the X.
The ruptured P2 aneurysm was dutifully coiled
Axial DYNA of this — stop to scroll through images
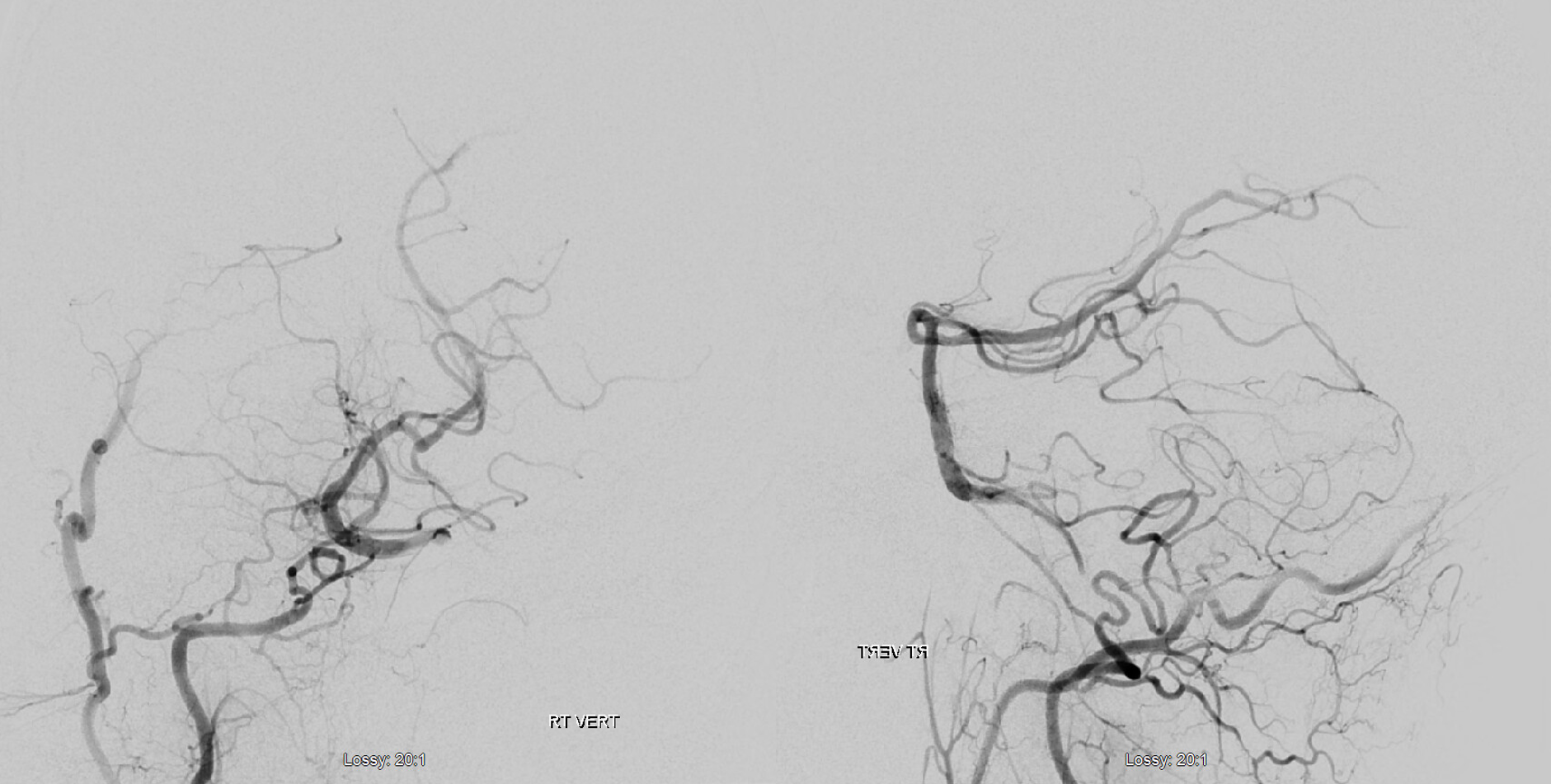
Another illustration, on opposite sides, in a patient with a complete circle of Willis, displayed to advantage by a generous injection.
“Normal” P1 segment origin from distal basilar (in a patient with duplicated AICAs)
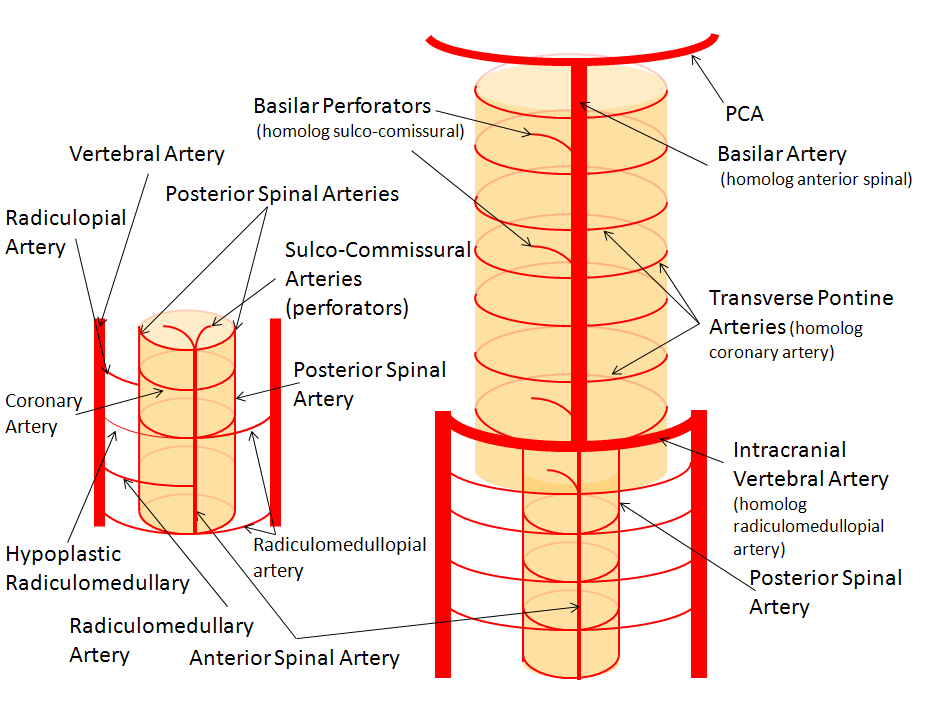
Conceptual homology of vertebrobasilar and spinal arterial anatomy, as it pertains to SCA and other vertebrobasilar arteries.
Lasjaunias and his colleagues view arterial system of the brainstem and cerebellum as a natural extension of the segmental arrangement found in the spinal cord.
The conceptual brilliance of this view allows one to understand all the myriad variations to which the basilar artery and its daughter vessels, such as the SCA, are subjected. In other words, if you consider the basilar artery to be a continuation of the anterior spinal artery, and its named branches and perforators as homologs of the coronary and sulco-comissural arteries (see Spinal Vascular Anatomy section), then the overall arrangement and its possible variations make perfect sense.
The following diagrams serve to illustrate this concept. On a personal note, I generally find anatomical diagrams to be at least somewhat wanting; when applied to the living body, they too often suffer from both rigidity and inconsistency, and almost universally fall short of the predictive potential for which their creation was originally intended. In this case, however, I believe that the genius of Lasjaunias (and supporting giants), may prove an exception. It is not, by any stretch, The Periodic Table, but some time investment into a bit of theoretical discussion is likely to produce major dividends.
Below is a diagram of cervical spinal vasculature (left), and brainstem vasculature (right), without the cerebellum.
The image on the LEFT represents cervical spinal cord arterial supply, which consists of the anterior spinal artery and a paired, loose network of posterolateral vessels known as the posterior spinal arteries, and which are conceptually represented here as contiguous vessels (which is at least mostly true in the cervical spine). The anterior and posterior spinal systems are connected by anastomoses running along the circumference of the cord, although known as “coronary” arteries, are conceptually quite clear. A number of perforating arteries into the substance of the cord exist; when arising from the anterior spinal artery and penetrating through the ventral cord sulcus, they are named “sulco-comissural” arteries. The entire spinal cord system is supplied via segmental radiculomedullary arteries, which connect the vertebral artery to the anterior spinal artery. In practice, as you know, the radiculomedullary and radiculopial arteries are fewer, and may arise from longitudinal vessels other than the vert. Radiculopial arteries are those which supply the posterior spinal system. Radiculomedullopial arteries are those which happen to supply both anterior and posterior spinal systems simultaneously, sometimes via a coronary artery or via separate connections. For a more complete discussion of spinal vasculature (essential, I believe), see Spinal Vascular Anatomy section, particularly Spinal Arterial Anatomy.
Now, lets add the brainstem to the spinal cord, and use existing arterial vascular networks to furnish its supply. Think of the brainstem as just a somewhat larger diameter biomass than the spinal cord, and things start to make sense. The unpaired basilar artery is a homolog of the equally unpaired anterior spinal artery. The intracranial vertebral arteries, although obliquely oriented, are essentially homologs of the radiculo-medullary arteries, inasmuch as they serve as transverse connections between the extraspinal vertebral system and the anterior spinal axis. The transverse pontine arteries are homologs of the coronary arteries. The basilar perforators are homologs of the sulco-comissural arteries. The posterior spinal arteries, in the superior cervical spine, are sometimes termed “Lateral spinal arteries”. This creates much unnecessary confusion, but the posterior spinal system and lateral spinal system are one and the same longitudinal arrangement.
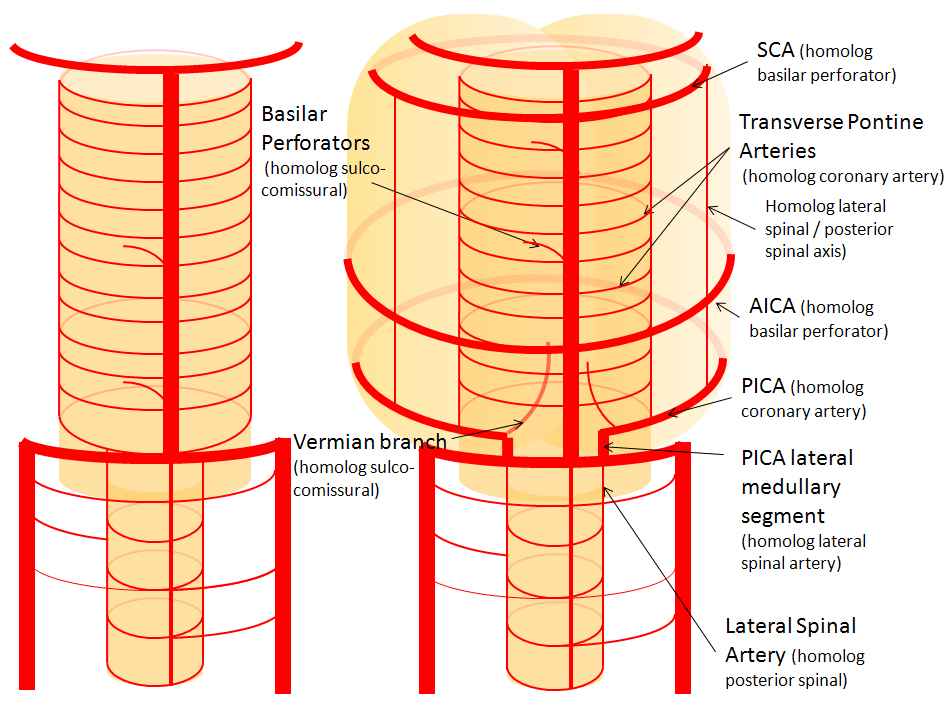
Now, add a cerebellum to the back of the brainstem, — again, simply more biomass — and use existing arteries to supply it. As the cerebellum develops, some of the transverse pontine perforators are recruited to capture the cerebellar hemispheric territory. Superiorly, this happens repatively consistently, and produces what is known as the Superior Cerebellar Artery. At the mid to lower basilar segment, a homologous enlarging channel is the AICA. At the bottom, the Posterior Inferior Cerebellar Artery (PICA) is the latest addition to cerebellar supply, Unlike AICA and SCA, it seems to arise from the lateral spinal system (yet nevertheless also a coronary artery homolog). The vermian arteries (of which only inferior is shown here) may be regarded as homologs of the sulco-comissural vessels.
There are many advantages to viewing the vertebrobasilar system in this way. All kinds of variants become quite predictable. For example, duplicated and triplicated SCAs and AICAs are simply persistence of adjacent transverse pontine (or midbrain) arteries in supply of the cerebellar hemisphere. AICAs arising higher or lower along the basilar are either results of dominance of higher or lower transverse arteries, or consequent to a relatively “short” basilar artery fusion. C1 origin of PICA reflects dominance of the C1 radiculopial artery, which via the C1 segment of the lateral medullary artery, gives rise to the PICA. The AICA-PICA balance in extent of cerebellar territory capture is a consequence of either anterior spinal (AICA) or lateral spinal (PICA) dominance. All of these cases are given angiographic illustrations below.
Basilar artery perforators
Classical disposition depicts AICA and SCA arising from the basilar artery, in addition to multiple short basilar perforators whose supply is limited to the brainstem. Understanding the embryology of vertebrobasilar circulation helps explain many variations seen in this pattern. As explained above, one can think of the basilar as a longitudinal channel, with multiple transverse channels. As the cerebellum develops, some of these transverse channels enlarge to capture the cortical territory of the cerebellum — thus becoming AICAs and SCAs. Others stay relatively small and are thus “limited” to brainstem supply. This simple concept explains ALL variations seen in the area. For example, duplicated SCAs and AICAs are just two adjacent perforators, which persisted in co-dominance of cerebellar supply. Dominant AICAs and PICAs and SCAs are simply variations in extent of cerebellar surface capture by one vessel, with corresponding dominance or hypoplasia of the others. “Larger” vs. smaller basilar perforators are but transverse channels which may capture some small segment of anterior cerebellar surface that is not taken by adjacent AICAs or SCAs.
Also important to recognize is the fact that brainstem perforators usually “arise” from SCA and AICA — or, rather, that AICA and SCA in fact “arise” from such perforators. Important, though very small and thus typically angiographically invisible, collaterals exist between these perforators. It is probable that extent of such collateral supply underlies the tremendous clinical recovery occasionally observed following brainstem infarcts.
Almost always, one or more “large” perforator is present between the AICA and the SCA — so consistently visualized, in fact, that it might almost deserve a unique name (we agreed to name it “ziggy” — or rather “Arteria ziggus basilarius” — until we found out that it already has a name — “Transverse Ponine Artery”). Occlusion of these transverse pontine arteries typically corresonds to ventral pontine infarction, and may lead to the ischemic cause of dreaded “locked in” syndrome.
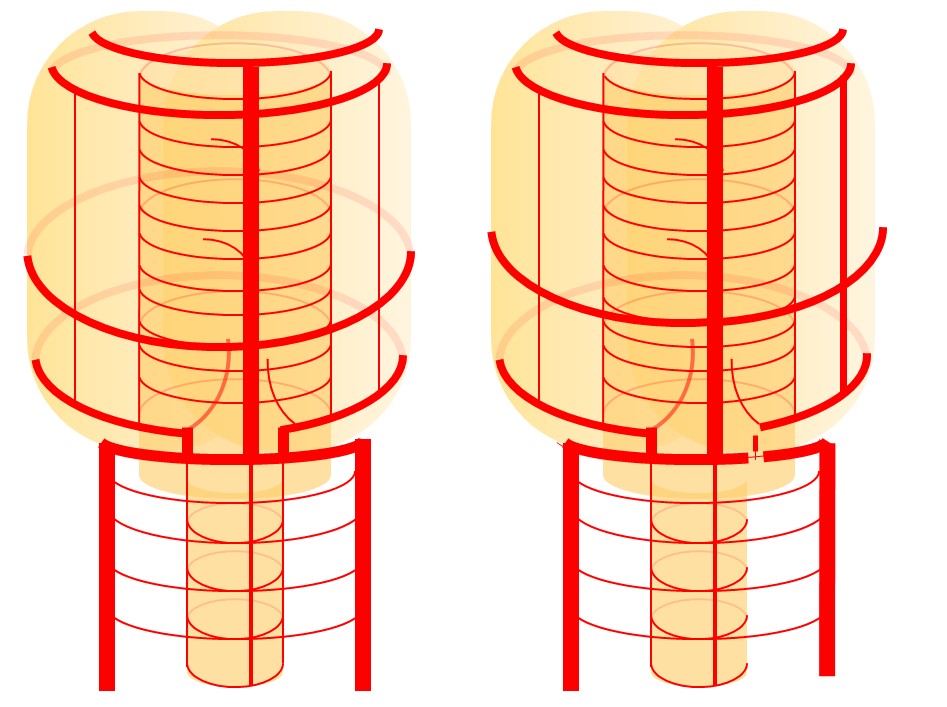
Diagram of the above concept. Image on left, with brainstem alone, shows schematic of vertebrobasilar system with numerous “transverse” perforators supplying the brainstem (transverse pontine arteries). Image on right, with cerebellum in place, depicts SCA, AICA, and PICA as perforators which enlarged to capture cerebellar hemispheric territory. This viewpoint allows for ready conceptualization of multiple SCA, AICA, and PICA variations observed within the vertebrobasilar system.
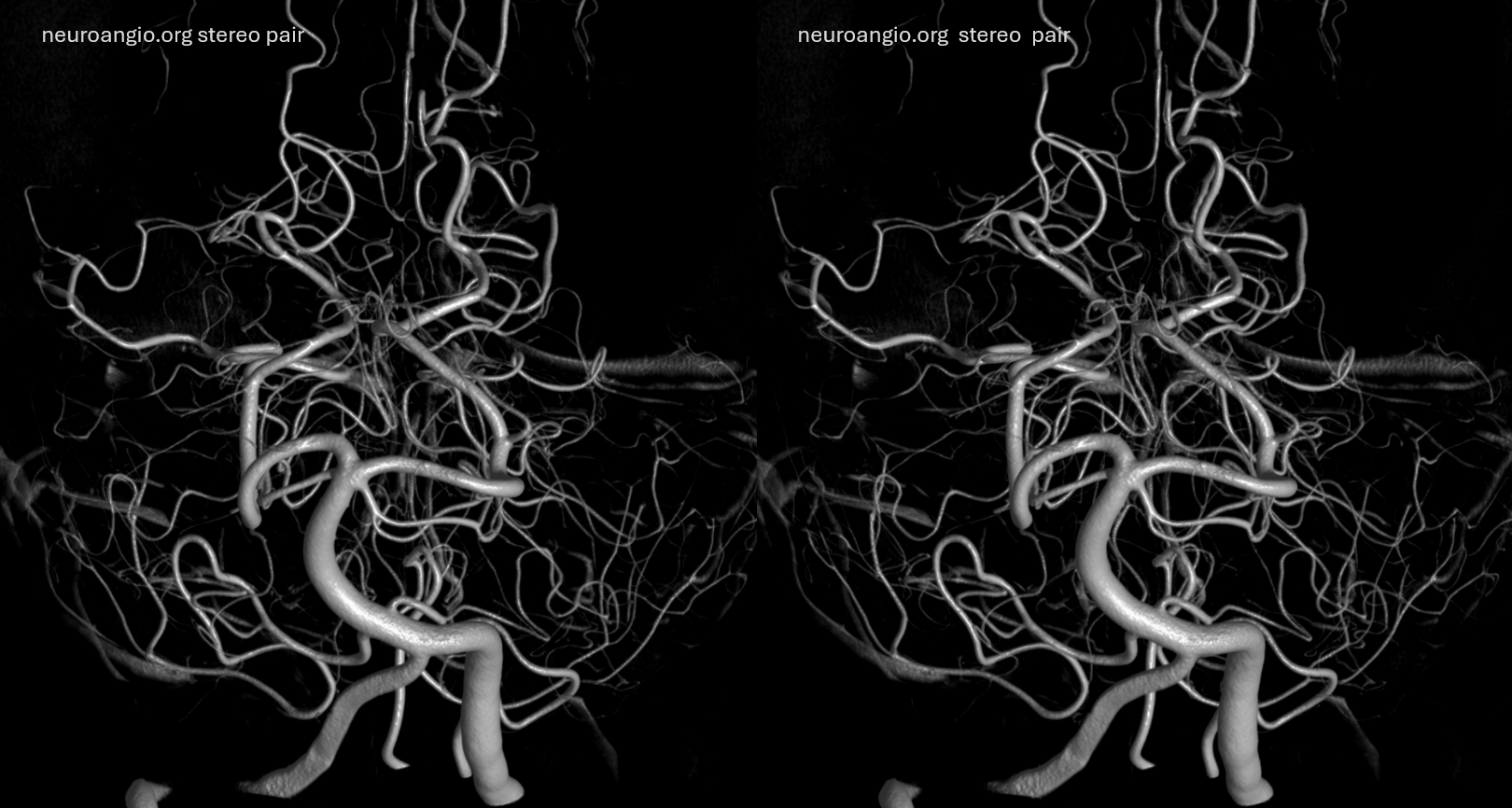
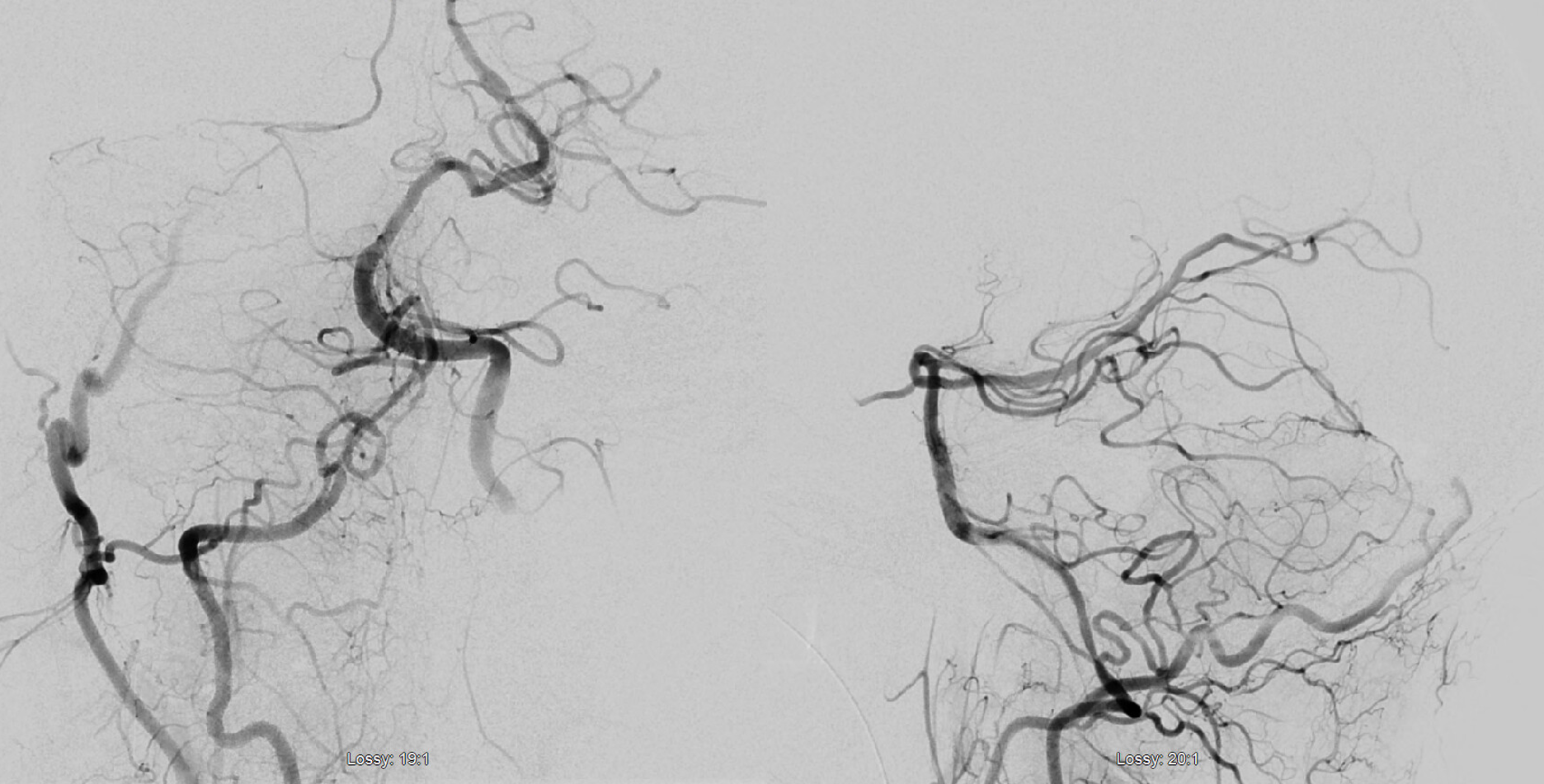
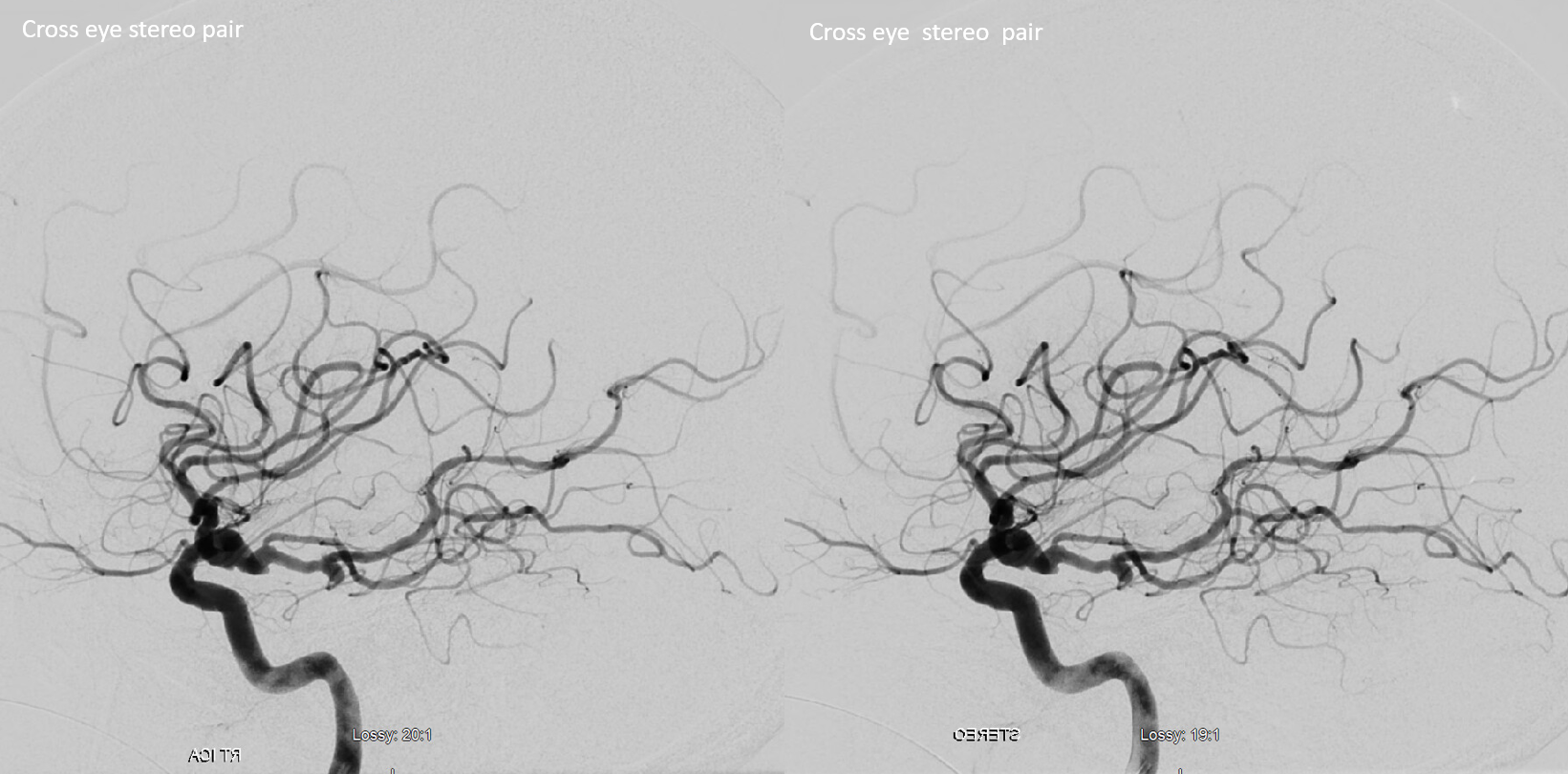
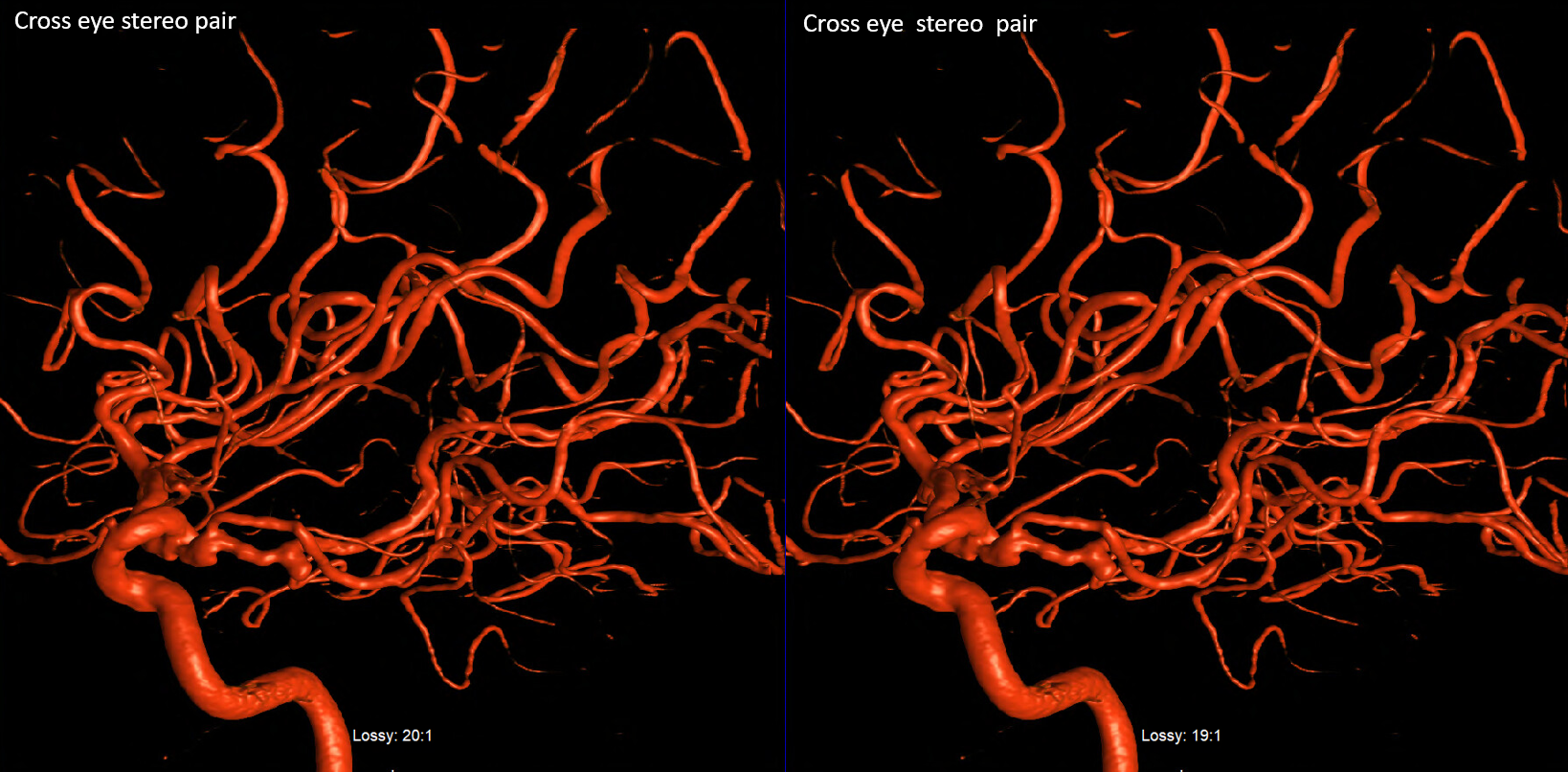
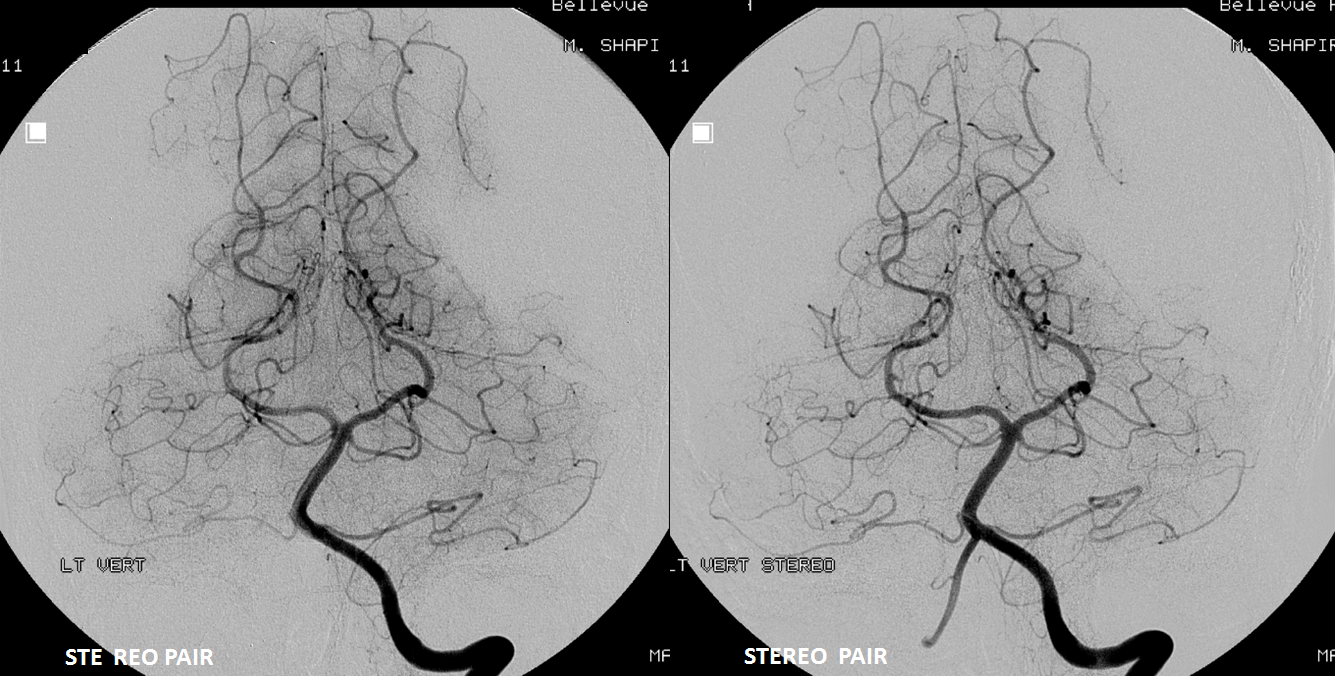
SCA duplication, stereo pair. Co-dominance of adjacent basilar perforators in capture of cerebellar territory results in SCA duplication, similar to situation sen with AICA. The SCA is unilaterally duplicated about 25% of time.
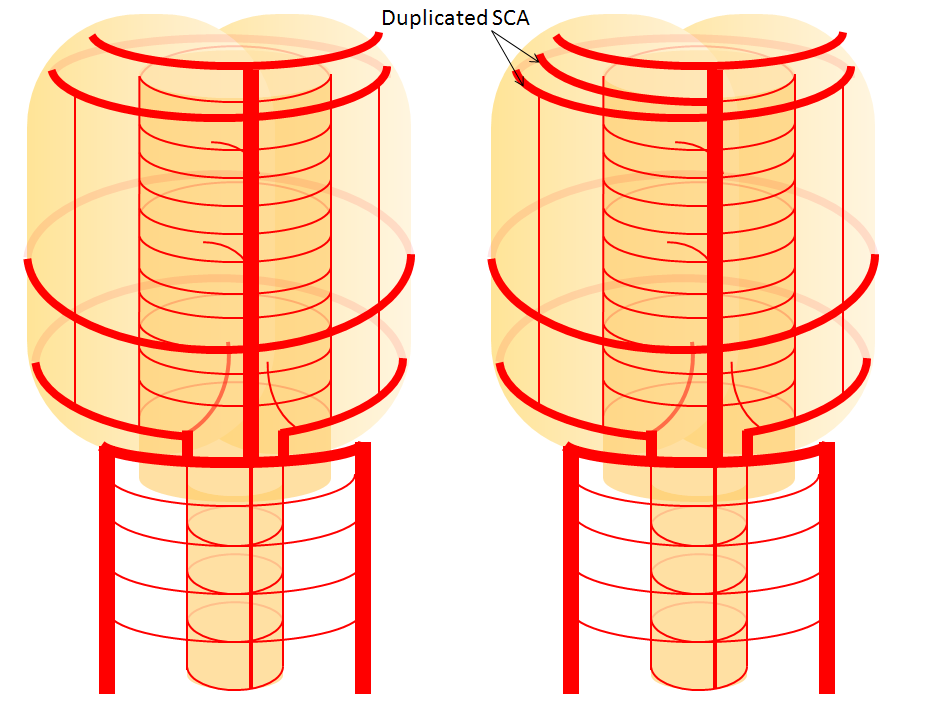
Diagram of the disposition (right image), as compared with the classical arrangement depicted on the left.
Superior Cerebellar Artery supply of PICA territory
Left vertebral artery injections (top images) demonstrate severe stenoses of the PICA segment (yellow), and PICA ostium itself (purple). The bottom images, in late arterial and capillary phases, show the hemispheric branch of the left SCA (red) extending inferomedially to reconstitute the territory of the incapacitated PICA.
Below is a diagram of the same adaptation
The above disposition need not come because of disease. As neuroangio emphasizes, everything is in balance. A given artery can only be understood in context of its relationship to other vessels with which it is in balance.
SCA – AICA-PICA balance
The overarching notion of “spectrum” in on display here. The important thing is territory of supply, not site of origin. How to put it all together? The right AICA (arrowheads) is small. Right PICA (dashed white arrow) supply is limited to vermis. What then supplies most of right cerebellar hemisphere — in this case, the SCA (white arrow) — note larger caliber of right SCA compared to left. On the left, the AICA is also hypoplastic (black arrowheads). The PICA (dashed black arrows) is dominant, and controls lateral hemisphere. Right SCA holds usual superior surface.
Artery of Wollschleger and Wollschleger
Piodural anastomosis between the SCA and posterior dura. Rare — much less prevalent than the PCA piodural branch known as artery of Davidoff-Schechter. Usually invisible angiographically when “normal”. In balance with other sources of the posterior meningeal artery. When pathologic contributes to various dural lesions — dural fistulas, meningiomas. Below is a rare example of one seen on DSA
Technique is of course important. its a small artery. also, its dural — which means that its vascular bed resistance will be higher than brain as a rule. which means that it will be seen in a later phase than brain arteries. thats why recurrent meningeal (Ophthalmic artery origin of the MMA) is best seen in late arterial or parenchymal brain phases of the ICA injection. Here its the same — see below: arterial phase (left) shows nothing. Parenchymal brain phase shows both the Davidoff-Schechter (PCA origin, solid arrow) and Wollschleger-Wollschleger arteries (SCA origin, projecting inferiorly, dashed arrows)